Single-cell multimodal analyses reveal epigenomic and transcriptomic basis for birth defects in maternal diabetes
Single-cell multimodal analyses reveal epigenomic and transcriptomic basis for birth defects in maternal diabetes"
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT Maternal diabetes mellitus is among the most frequent environmental contributors to congenital birth defects, including heart defects and craniofacial anomalies, yet the cell types
affected and mechanisms of disruption are largely unknown. Here, using multimodal single-cell analyses, we show that maternal diabetes affects the epigenomic landscape of specific subsets of
cardiac and craniofacial progenitors during embryogenesis. A previously unrecognized cardiac progenitor subpopulation expressing the homeodomain-containing protein ALX3 showed prominent
chromatin accessibility changes and acquired a more posterior identity. Similarly, a subpopulation of neural crest-derived cells in the second pharyngeal arch, which contributes to
craniofacial structures, displayed abnormalities in the epigenetic landscape and axial patterning defects. Chromatin accessibility changes in both populations were associated with increased
retinoic acid signaling, known to establish anterior–posterior identity. This work highlights how an environmental insult can have highly selective epigenomic consequences on discrete cell
types leading to developmental patterning defects. Access through your institution Buy or subscribe This is a preview of subscription content, access via your institution ACCESS OPTIONS
Access through your institution Subscribe to this journal Receive 12 digital issues and online access to articles $119.00 per year only $9.92 per issue Learn more Buy this article * Purchase
on SpringerLink * Instant access to full article PDF Buy now Prices may be subject to local taxes which are calculated during checkout ADDITIONAL ACCESS OPTIONS: * Log in * Learn about
institutional subscriptions * Read our FAQs * Contact customer support SIMILAR CONTENT BEING VIEWED BY OTHERS SINGLE-CELL TRANSCRIPTOMIC PROFILING UNVEILS DYSREGULATION OF CARDIAC PROGENITOR
CELLS AND CARDIOMYOCYTES IN A MOUSE MODEL OF MATERNAL HYPERGLYCEMIA Article Open access 15 August 2022 SINGLE-CELL CHROMATIN PROFILING OF THE PRIMITIVE GUT TUBE REVEALS REGULATORY DYNAMICS
UNDERLYING LINEAGE FATE DECISIONS Article Open access 26 May 2022 SINGLE-CELL RNA SEQUENCING IDENTIFIES CXADR AS A FATE DETERMINANT OF THE PLACENTAL EXCHANGE SURFACE Article Open access 02
January 2025 DATA AVAILABILITY Data are available in the main text and the supplementary materials. All the sequencing data have been deposited in NCBI’s Gene Expression Omnibus and are
accessible through GEO series accession number GSE198905. Source data are provided with this paper. CODE AVAILABILITY All codes are available on GitHub
(https://github.com/SrivastavaLab-Gladstone/Nishino_DM_2022). REFERENCES * Hutson, M. R. & Kirby, M. L. Neural crest and cardiovascular development: a 20-year perspective. _Birth Defects
Res. C_ 69, 2–13 (2003). Article CAS Google Scholar * Srivastava, D. Making or breaking the heart: from lineage determination to morphogenesis. _Cell_ 126, 1037–1048 (2006). Article CAS
PubMed Google Scholar * Ogurtsova, K. et al. IDF Diabetes Atlas: global estimates for the prevalence of diabetes for 2015 and 2040. _Diabetes Res. Clin. Pract._ 128, 40–50 (2017).
Article CAS PubMed Google Scholar * Lammer, E. J. et al. Retinoic acid embryopathy. _N. Engl. J. Med._ 313, 837–841 (1985). Article CAS PubMed Google Scholar * Langston, A. W. &
Gudas, L. J. Retinoic acid and homeobox gene regulation. _Curr. Opin. Genet. Dev._ 4, 550–555 (1994). Article CAS PubMed Google Scholar * Waxman, J. S. & Yelon, D. Increased Hox
activity mimics the teratogenic effects of excess retinoic acid signaling. _Dev. Dyn._ 238, 1207–1213 (2009). Article CAS PubMed PubMed Central Google Scholar * Basu, M. & Garg, V.
Maternal hyperglycemia and fetal cardiac development: clinical impact and underlying mechanisms. _Birth Defects Res. A_ 110, 1504–1516 (2018). Article CAS Google Scholar * Helle, E. &
Priest, J. R. Maternal obesity and diabetes mellitus as risk factors for congenital heart disease in the offspring. _J. Am. Heart Assoc._ 9, e011541 (2020). Article CAS PubMed PubMed
Central Google Scholar * Jovanovic, L. et al. The Diabetes in Early Pregnancy Study: β-hydroxybutyrate levels in type 1 diabetic pregnancy compared with normal pregnancy. NICHD-Diabetes in
Early Pregnancy Study Group (DIEP). National Institute of Child Health and Development. _Diabetes Care_ 21, 1978–1984 (1998). Article CAS PubMed Google Scholar * Xie, Z. et al.
Metabolic regulation of gene expression by histone lysine β-hydroxybutyrylation. _Mol. Cell_ 62, 194–206 (2016). Article CAS PubMed PubMed Central Google Scholar * Moazzen, H. et al.
N-Acetylcysteine prevents congenital heart defects induced by pregestational diabetes. _Cardiovasc. Diabetol._ 13, 46 (2014). Article PubMed PubMed Central Google Scholar * Yang, P. et
al. Maternal hyperglycemia activates an ASK1–FoxO3a–caspase 8 pathway that leads to embryonic neural tube defects. _Sci. Signal._ 6, ra74 (2013). Article PubMed PubMed Central Google
Scholar * Hrubec, T. C., Prater, M. R., Toops, K. A. & Holladay, S. D. Reduction in diabetes-induced craniofacial defects by maternal immune stimulation. _Birth Defects Res. B_ 77, 1–9
(2006). Article CAS Google Scholar * Hao, Y. et al. Integrated analysis of multimodal single-cell data. _Cell_ 184, 3573–3587 e3529 (2021). Article CAS PubMed PubMed Central Google
Scholar * Granja, J. M. et al. ArchR is a scalable software package for integrative single-cell chromatin accessibility analysis. _Nat. Genet._ 53, 403–411 (2021). Article CAS PubMed
PubMed Central Google Scholar * Heinz, S. et al. Simple combinations of lineage-determining transcription factors prime cis-regulatory elements required for macrophage and B cell
identities. _Mol. Cell_ 38, 576–589 (2010). Article CAS PubMed PubMed Central Google Scholar * Simoes-Costa, M. & Bronner, M. E. Establishing neural crest identity: a gene
regulatory recipe. _Development_ 142, 242–257 (2015). Article CAS PubMed PubMed Central Google Scholar * Parker, H. J., Pushel, I. & Krumlauf, R. Coupling the roles of Hox genes to
regulatory networks patterning cranial neural crest. _Dev. Biol._ 444, S67–S78 (2018). Article CAS PubMed Google Scholar * Dooley, C. M. et al. The gene regulatory basis of genetic
compensation during neural crest induction. _PLoS Genet._ 15, e1008213 (2019). Article CAS PubMed PubMed Central Google Scholar * Rothstein, M. & Simoes-Costa, M. Heterodimerization
of TFAP2 pioneer factors drives epigenomic remodeling during neural crest specification. _Genome Res._ 30, 35–48 (2020). Article CAS PubMed PubMed Central Google Scholar * Fan, X. et
al. TWIST1 and chromatin regulatory proteins interact to guide neural crest cell differentiation. _eLife_ https://doi.org/10.7554/eLife.62873 (2021). * Soldatov, R. et al. Spatiotemporal
structure of cell fate decisions in murine neural crest. _Science_ https://doi.org/10.1126/science.aas9536 (2019). * Akerberg, B. N. et al. A reference map of murine cardiac transcription
factor chromatin occupancy identifies dynamic and conserved enhancers. _Nat. Commun._ 10, 4907 (2019). Article PubMed PubMed Central Google Scholar * Prall, O. W. et al. An
Nkx2-5/Bmp2/Smad1 negative feedback loop controls heart progenitor specification and proliferation. _Cell_ 128, 947–959 (2007). Article CAS PubMed PubMed Central Google Scholar *
Schussler, O. et al. Cardiac neural crest cells: their rhombomeric specification, migration, and association with heart and great vessel anomalies. _Cell Mol. Neurobiol._ 41, 403–429 (2021).
Article CAS PubMed Google Scholar * Rada-Iglesias, A. et al. Epigenomic annotation of enhancers predicts transcriptional regulators of human neural crest. _Cell Stem Cell_ 11, 633–648
(2012). Article CAS PubMed PubMed Central Google Scholar * Maconochie, M. et al. Regulation of Hoxa2 in cranial neural crest cells involves members of the AP-2 family. _Development_
126, 1483–1494 (1999). Article CAS PubMed Google Scholar * Kutejova, E., Engist, B., Mallo, M., Kanzler, B. & Bobola, N. Hoxa2 downregulates Six2 in the neural crest-derived
mesenchyme. _Development_ 132, 469–478 (2005). Article CAS PubMed Google Scholar * Van Otterloo, E., Li, H., Jones, K. L. & Williams, T. AP-2α and AP-2β cooperatively orchestrate
homeobox gene expression during branchial arch patterning. _Development_ 145, dev157438 (2018). PubMed PubMed Central Google Scholar * Kirilenko, P. et al. Transient activation of meox1
is an early component of the gene regulatory network downstream of hoxa2. _Mol. Cell. Biol._ 31, 1301–1308 (2011). Article CAS PubMed PubMed Central Google Scholar * Alexanian, M. et
al. A transcriptional switch governs fibroblast activation in heart disease. _Nature_ 595, 438–443 (2021). Article CAS PubMed PubMed Central Google Scholar * Kruzynska-Frejtag, A. et
al. Periostin is expressed within the developing teeth at the sites of epithelial–mesenchymal interaction. _Dev. Dyn._ 229, 857–868 (2004). Article CAS PubMed Google Scholar * Verzi, M.
P., McCulley, D. J., De Val, S., Dodou, E. & Black, B. L. The right ventricle, outflow tract, and ventricular septum comprise a restricted expression domain within the secondary/anterior
heart field. _Dev. Biol._ 287, 134–145 (2005). Article CAS PubMed Google Scholar * de Soysa, T. Y. et al. Single-cell analysis of cardiogenesis reveals basis for organ-level
developmental defects. _Nature_ 572, 120–124 (2019). Article PubMed PubMed Central Google Scholar * Beverdam, A., Brouwer, A., Reijnen, M., Korving, J. & Meijlink, F. Severe nasal
clefting and abnormal embryonic apoptosis in Alx3/Alx4 double mutant mice. _Development_ 128, 3975–3986 (2001). Article CAS PubMed Google Scholar * Basson, M. A. Signaling in cell
differentiation and morphogenesis. _Cold Spring Harb. Perspect. Biol_. https://doi.org/10.1101/cshperspect.a008151 (2012). * Stefanovic, S. et al. Hox-dependent coordination of mouse cardiac
progenitor cell patterning and differentiation. _eLife_ https://doi.org/10.7554/eLife.55124 (2020). * Pineault, N., Helgason, C. D., Lawrence, H. J. & Humphries, R. K. Differential
expression of Hox, Meis1, and Pbx1 genes in primitive cells throughout murine hematopoietic ontogeny. _Exp. Hematol._ 30, 49–57 (2002). Article CAS PubMed Google Scholar * Arrington, C.
B., Dowse, B. R., Bleyl, S. B. & Bowles, N. E. Non-synonymous variants in pre-B cell leukemia homeobox (PBX) genes are associated with congenital heart defects. _Eur. J. Med. Genet._ 55,
235–237 (2012). Article PubMed PubMed Central Google Scholar * Stennard, F. A. et al. Murine T-box transcription factor Tbx20 acts as a repressor during heart development, and is
essential for adult heart integrity, function and adaptation. _Development_ 132, 2451–2462 (2005). Article CAS PubMed Google Scholar * Xavier-Neto, J. et al. A retinoic acid-inducible
transgenic marker of sino-atrial development in the mouse heart. _Development_ 126, 2677–2687 (1999). Article CAS PubMed Google Scholar * Langfelder, P. & Horvath, S. WGCNA: an R
package for weighted correlation network analysis. _BMC Bioinf._ 9, 559 (2008). Article Google Scholar * Furman, B. L. Streptozotocin-induced diabetic models in mice and rats. _Curr.
Protoc._ 1, e78 (2021). Article CAS PubMed Google Scholar * Korsunsky, I. et al. Fast, sensitive and accurate integration of single-cell data with Harmony. _Nat. Methods_ 16, 1289–1296
(2019). Article CAS PubMed PubMed Central Google Scholar * Chen, E. Y. et al. Enrichr: interactive and collaborative HTML5 gene list enrichment analysis tool. _BMC Bioinf._ 14, 128
(2013). Article Google Scholar * Sarropoulos, I. et al. Developmental and evolutionary dynamics of cis-regulatory elements in mouse cerebellar cells. _Science_
https://doi.org/10.1126/science.abg4696 (2021). * Schep, A. N., Wu, B., Buenrostro, J. D. & Greenleaf, W. J. chromVAR: inferring transcription-factor-associated accessibility from
single-cell epigenomic data. _Nat. Methods_ 14, 975–978 (2017). Article CAS PubMed PubMed Central Google Scholar * Miller, S. A. et al. LSD1 and aberrant DNA methylation mediate
persistence of enteroendocrine progenitors that support BRAF-mutant colorectal cancer. _Cancer Res._ 81, 3791–3805 (2021). * Zhu, L. J. et al. ChIPpeakAnno: a Bioconductor package to
annotate ChIP-seq and ChIP-chip data. _BMC Bioinf._ 11, 237 (2010). Article Google Scholar * Yu, G., Wang, L. G. & He, Q. Y. ChIPseeker: an R/Bioconductor package for ChIP peak
annotation, comparison and visualization. _Bioinformatics_ 31, 2382–2383 (2015). Article CAS PubMed Google Scholar * McLean, C. Y. et al. GREAT improves functional interpretation of
_cis_-regulatory regions. _Nat. Biotechnol._ 28, 495–501 (2010). Article CAS PubMed PubMed Central Google Scholar * Tanigawa, Y., Dyer, E. S. & Bejerano, G. Which TF is functionally
important in your open chromatin data? _PLoS Comput Biol._ 18, e1010378 (2022). * Zolfaghari, R. et al. CYP26A1 gene promoter is a useful tool for reporting RAR-mediated retinoid activity.
_Anal. Biochem._ 577, 98–109 (2019). Article CAS PubMed PubMed Central Google Scholar * Zolfaghari, R. & Ross, A. C. Hepatocyte nuclear factor 4α (HNF4α) in coordination with
retinoic acid receptors increases all-_trans_-retinoic acid-dependent CYP26A1 gene expression in HepG2 human hepatocytes. _J. Cell. Biochem._ 115, 1740–1751 (2014). Article CAS PubMed
PubMed Central Google Scholar Download references ACKNOWLEDGEMENTS We thank members of the Srivastava laboratory for discussion and feedback; B. Taylor from Gladstone Institutes for
editorial and graphics assistance; G. Maki from Gladstone Institutes for graphics assistance; and K. Claiborn from Gladstone Institutes for editorial review. We acknowledge the Center for
Advanced Technology (CAT) for sequencing; the Gladstone Histology and Light Microscopy Core for their technical support; and the Gladstone Animal Facility for support with mouse colonies.
Figures 1a and 5g, Extended Data Fig. 1a and Supplementary Fig. 1a were created with BioRender.com. National Institutes of Health/NHLBI grant P01 HL146366, R01 HL057181, R01 HL015100, R01
HL127240, Roddenberry Foundation, L.K. Whittier Foundation, Dario and Irina Sattui, Younger Family Fund, and Additional Ventures to D.S. The Japan Society for the Promotion of Science
Overseas Research Fellowship to T.N. Additional Ventures to S.S.R. American Heart Association Postdoctoral Fellowship (#899270) to B.J.v.S. National Institutes of Health grant K08 HL157700,
Sarnoff Cardiovascular Research Foundation, Frank A. Campini Foundation and Michael Antonov Charitable Foundation to A. Padmanabhan. AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * Gladstone
Institutes, San Francisco, CA, USA Tomohiro Nishino, Sanjeev S. Ranade, Angelo Pelonero, Benjamin J. van Soldt, Lin Ye, Michael Alexanian, Frances Koback, Yu Huang, Langley Grace Wallace,
Nandhini Sadagopan, Adrienne Lam, Lyandysha V. Zholudeva, Feiya Li, Arun Padmanabhan, Reuben Thomas, Joke G. van Bemmel, Casey A. Gifford, Mauro W. Costa & Deepak Srivastava *
Roddenberry Center for Stem Cell Biology and Medicine at Gladstone, San Francisco, CA, USA Tomohiro Nishino, Sanjeev S. Ranade, Angelo Pelonero, Benjamin J. van Soldt, Lin Ye, Michael
Alexanian, Frances Koback, Yu Huang, Langley Grace Wallace, Nandhini Sadagopan, Adrienne Lam, Lyandysha V. Zholudeva, Feiya Li, Arun Padmanabhan, Joke G. van Bemmel, Casey A. Gifford, Mauro
W. Costa & Deepak Srivastava * Department of Pediatrics, University of California, San Francisco, San Francisco, CA, USA Michael Alexanian & Deepak Srivastava * Division of
Cardiology, Department of Medicine, University of California, San Francisco, San Francisco, CA, USA Nandhini Sadagopan & Arun Padmanabhan * Chan Zuckerberg Biohub, San Francisco, CA, USA
Arun Padmanabhan * Department of Biochemistry and Biophysics, University of California, San Francisco, San Francisco, CA, USA Deepak Srivastava Authors * Tomohiro Nishino View author
publications You can also search for this author inPubMed Google Scholar * Sanjeev S. Ranade View author publications You can also search for this author inPubMed Google Scholar * Angelo
Pelonero View author publications You can also search for this author inPubMed Google Scholar * Benjamin J. van Soldt View author publications You can also search for this author inPubMed
Google Scholar * Lin Ye View author publications You can also search for this author inPubMed Google Scholar * Michael Alexanian View author publications You can also search for this author
inPubMed Google Scholar * Frances Koback View author publications You can also search for this author inPubMed Google Scholar * Yu Huang View author publications You can also search for this
author inPubMed Google Scholar * Langley Grace Wallace View author publications You can also search for this author inPubMed Google Scholar * Nandhini Sadagopan View author publications You
can also search for this author inPubMed Google Scholar * Adrienne Lam View author publications You can also search for this author inPubMed Google Scholar * Lyandysha V. Zholudeva View
author publications You can also search for this author inPubMed Google Scholar * Feiya Li View author publications You can also search for this author inPubMed Google Scholar * Arun
Padmanabhan View author publications You can also search for this author inPubMed Google Scholar * Reuben Thomas View author publications You can also search for this author inPubMed Google
Scholar * Joke G. van Bemmel View author publications You can also search for this author inPubMed Google Scholar * Casey A. Gifford View author publications You can also search for this
author inPubMed Google Scholar * Mauro W. Costa View author publications You can also search for this author inPubMed Google Scholar * Deepak Srivastava View author publications You can also
search for this author inPubMed Google Scholar CONTRIBUTIONS T.N. and D.S. conceived and directed the study. T.N. and Y.H. performed animal work. T.N., L.G.W. and S.S.R. collected heart
tissues and isolated single cells for subsequent scRNA-seq and scATAC-seq. T.N., S.S.R., A. Pelonero, B.J.v.S. and F.K. analyzed scRNA-seq and scATAT-seq and developed computational methods.
T.N., B.J.v.S., L.V.Z. and F.L. performed RNA in situ hybridization and subsequent tissue clearing and imaging. T.N., L.Y., N.S., A.L., A. Padmanabhan and M.W.C designed, performed and
analyzed luciferase assay and mouse lineage trace experiment. T.N., S.S.R., M.A., A. Pelonero, J.G.v.B, C.A.G., M.W.C. and D.S. interpreted the data. R.T. reviewed statistical methods. T.N.,
M.W.C. and D.S. wrote the manuscript with contributions of M.A. CORRESPONDING AUTHOR Correspondence to Deepak Srivastava. ETHICS DECLARATIONS COMPETING INTERESTS D.S. is a scientific
co-founder, shareholder and director of Tenaya Therapeutics. The remaining authors declare no competing interests. PEER REVIEW PEER REVIEW INFORMATION _Nature Cardiovascular Research_ thanks
Professor Hiroki Kurihara, and the other, anonymous, reviewer for their contribution to the peer review of this work. Primary Handling Editor: Vesna Todorovic, in collaboration with the
_Nature Cardiovascular Research_ team. ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional
affiliations. EXTENDED DATA EXTENDED DATA FIG. 1 HISTOLOGICAL AND MICRO-CT VALIDATION OF THE MATERNAL DIABETES MODEL. (A) The design of the _in vivo_ maternal diabetic model experiment.
After administration of either VEH or STZ, females in the STZ group with confirmed diabetes were mated with normoglycemic males, and heart samples at embryonic day 18.5 (E18.5) or postnatal
day 0 (P0) were collected for histological examination. (B) Representative micro-CT images of the heart phenotypes detected in the diabetic model. The prevalence of each malformation is
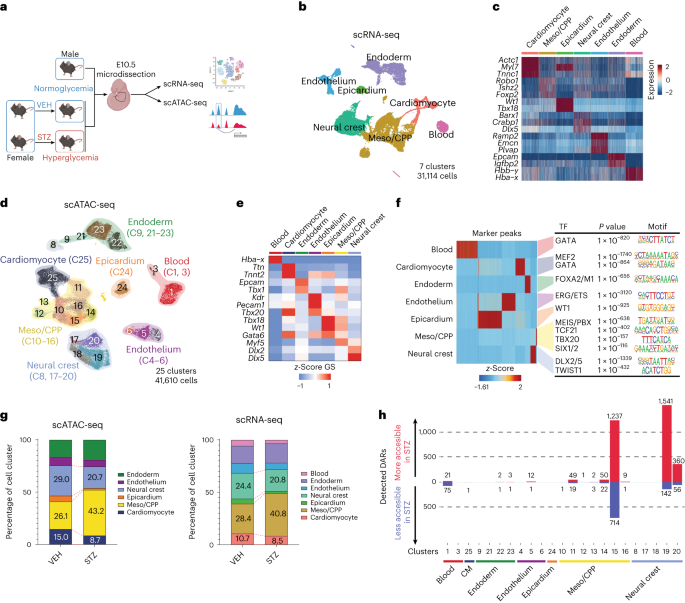
shown in Supplementary Table 1. The scale bar represents 500 µm. EXTENDED DATA FIG. 2 SINGLE CELL MULTIMODAL ANALYSIS OF CARDIO-PHARYNGEAL REGION IN MATERNAL DIABETES. (A) Representative
image of E10.5 embryo with detailed micro-dissected region used for scRNA/scATAC-seq experiment. Scale bar represents 1 mm. (B) scRNA-seq (left) or scATAC-seq (right) UMAP presentation
colored by conditions. Different colors overlayed delineate cell type cluster annotations. (C) Expression patterns of representative cell type specific marker genes plotted on UMAP space
shown in Fig. 1b. (D) Heatmap of marker gene scores per cluster of scATAC-seq. (E) Heatmap of Jaccard indices calculated between scRNA-seq and scATAC-seq after integration. Values range from
0 to 1 (higher value represents closer annotation matching between the two modalities). (F) Genomic distribution of all the called peaks color coded by the genomic location as shown. Total
called peaks = 492,330. (G) Distribution of all called peaks based on the distance from transcription start sites. EXTENDED DATA FIG. 3 MATERNAL DIABETES DYSREGULATES EPIGENOMIC LANDSCAPE OF
NEURAL CREST CELLS IN PHARYNGEAL ARCHES 4 AND 6. (A) Population distribution by sub-cell-type normalized to total number of cells per sample in neural crest cell subset data of scRNA-seq.
Numbers inside the barplot represent the percentage of cell types of the total cell number. Statistics performed by permutation test in scRNA-seq data, comparing STZ vs. VEH, for NC-prog,
FDR < 0.001, Log2FD = 1.88; for SMC-prog, FDR < 0.001, Log2FD = −0.33. (B) Heatmap of Jaccard indices calculated between neural crest cell scRNA-seq and scATAC-seq cell annotations
after integration. Values range from 0 to 1 (the higher value represents closer annotation matching between those two modalities). (C) MA plot of DARs in PA3/4/6 population between VEH and
STZ. Red dots represent the more accessible (open) (FDR < = 0.05 & Log2FC > = 1) and blue dots represent less accessible (closed) DARs in STZ (FDR < = 0.05 & Log2FC < =
−1). (D) Enriched TF binding motifs in more accessible (left) or less accessible (right) DARs in STZ vs. VEH within the PA3/4/6 population. (E) scATAC-seq UMAP representation of neural crest
cell C20 subset population colored by clusters (PA3 – dark red; PA4/6 – dark blue). (F) Heatmap of Gene Scores (GS) of curated marker genes based on scRNA-seq data for PA3 and PA4/6 neural
crest. Scale indicates z-scored GS values. (G) MA plot of DARs between VEH and STZ in PA3 population (left) and PA4/6 population (right). Red dots represent the more accessible (open) (FDR
< = 0.05 & Log2FC > = 1) and blue dots represent less accessible (closed) DARs in STZ (FDR < = 0.05 & Log2FC < = −1). (H) Enriched TF binding motifs in more accessible
(left) and less accessible (right) DARs in STZ in PA4/6 population. NC-prog, neural crest cell progenitors; PA2, pharyngeal arch 2; PA3, pharyngeal arch 3; PA4/6, pharyngeal arch 4/6; SMC,
smooth muscle cells; SMC-prog, smooth muscle cell progenitors. Source data EXTENDED DATA FIG. 4 MATERNAL DIABETES DYSREGULATES EPIGENOMIC AND TRANSCRIPTIONAL LANDSCAPE ASSOCIATED WITH CELL
DIFFERENTIATION AND PATTERNING IN PHARYNGEAL ARCH 2 NEURAL CREST. (A) Violin plot of _Tfap2a_ expression levels in cluster 0 of UMAP in Fig. 2i across 3 VEH and 3 STZ embryos (Wilcoxon Rank
Sum test). (B) Expression of _Nr2f1_ mRNA on UMAP space for PA2 neural crest cells (VEH – left top; STZ – left bottom). Scale bar indicates z-scored expression values. Violin plot of _Nr2f1_
expression levels in cluster 0 of UMAP in Fig. 2i (right) (Wilcoxon Rank Sum test). (C) Expression of indicated genes on UMAP space for PA2 neural crest cells. Scale bar indicates z-scored
expression values. (D) Violin plots of _Dlx5_ (left) and _Dlx6_ (right) expression levels in cluster 0 of UMAP in Fig. 2i (Wilcoxon Rank Sum test). (E) Enriched GO terms in detected DARs in
PA2 population using GREAT analysis. (F) Enriched GO terms in detected DARs in PA3/4/6 population using GREAT analysis. EXTENDED DATA FIG. 5 IDENTIFICATION OF DISTINCT SUBSETS OF AHF
PROGENITORS. (A) scRNA-seq UMAP representation of mesodermal population (‘Meso/CPP’, ‘Cardiomyocyte’, or ‘Epicardium’ in Fig. 1b) colored by conditions (VEH – blue; STZ – light red). (B)
Heatmap of Jaccard indices between mesoderm cell scRNA-seq and scATAC-seq annotations after integration. Values range from 0 to 1 (the higher value represents closer annotation matching
between those two modalities). CM_V, ventricular cardiomyocyte; CM_AVC, atrioventricular canal cardiomyocyte; CM_A, atrial cardiomyocyte; CM_SV, sinus venosus cardiomyocyte; CM_OFT, outflow
tract cardiomyocyte; pSHF1/2, posterior second heart field 1/2; EndoMT, endothelial mesenchymal transition; EpiC, Epicardium; AHF1/2, anterior heart field 1/2; PharyngealMeso, pharyngeal
mesoderm; ParaxialMeso1/2, paraxial mesoderm 1/2; BrM, branchiomeric muscle. (C) Expression pattern of _Hand2_ (left) and _Rgs5 (right)_ on UMAP space. AHF1 and 2 are circled in red. Scale
bar indicates z-scored expression values. EXTENDED DATA FIG. 6 _ALX3_POS CELLS ARE A DISTINCT SUBSET OF THE AHF POPULATION. (A) Representative images from RNA _in situ_ hybridization for
_Armh4_ (green) and _Alx3_ (red) in an E10.5 embryo from VEH treated female. The scale bar represents 500 µm. (B) Representative images from whole mount RNA _in situ_ hybridization of E10.5
embryos using light sheet microscopy. _Armh4_ (green) and _Alx3_ (red) expression is shown from the dorsal view (D – left) and the right oblique view (O – right). A white bracket (left)
highlights the anterior part of _Alx3_Pos cells. A white dotted oval (right) highlights the _Alx3_Pos cell streak on left side of the embryo from outflow tract (OFT) towards the
posterolateral region. Still images were extracted from Supplementary video 2. Scale bar represents 100 µm. PA2, pharyngeal arch 2; BW, body wall. (C) The distribution of _Alx3_ positive
cells by scRNA-seq between E7.75 and E9.25. (D) scRNA-seq UMAP of cardiac progenitor cells at E9.25 from the same data as (C) color coded by cell type annotation. AHF, anterior heart field;
BM progenitors, branchiomeric muscle progenitors; pSHF, posterior second heart field. (E) Expression of _Alx3_ on the same UMAP as (D). (F) Heatmap of differentially expressed genes (DEGs)
between _Alx3_Neg AHF and _Alx3_Pos AHF at E9.25. All detected DEGs that attained adjusted p-val < 0.05 and Log2FC > 0.25 are shown. Top GO terms enriched in upregulated or
downregulated DEGs are shown with representative genes composing each GO (Fisher’s exact test, corrected for multiple testing using the Benjamini-Hochberg method). Scale bar indicates
z-scored expression values. (G) Heatmap presentation of DEGs between AHF1 and AHF2 at E10.5 using only VEH cells in the scRNA-seq data. All detected DEGs that attained adjusted p-val <
0.05 and Log2FC > 0.25 are shown. Top GO terms enriched in upregulated or downregulated DEGs are shown with representative genes composing each GO (Fisher’s exact test, corrected for
multiple testing using the Benjamini-Hochberg method). Scale bar indicates z-scored expression values. (H) Venn diagram representing the intersect between DEGs shown in (F-G). EXTENDED DATA
FIG. 7 PGDM DISRUPTS ANTERIOR-POSTERIOR PATTERNING IN AHF2. (A) Enriched GO terms in VEH vs. STZ DARs in AHF2 population using GREAT analysis. (B) Heatmap of marker genes of each of three
subclusters found in _Alx3_Pos AHF2. These marker genes were detected using only VEH-treated _Alx3_Pos AHF2 cells (left). All marker genes that attained an adjusted p-val < 0.05 and
Log2FC > 0.25 are shown. Scale bar indicates z-scored expression values. Top GO terms enriched in marker genes for each sub cluster with statistical information and representative maker
genes to corresponding GO term are shown (Fisher’s exact test, corrected for multiple testing using the Benjamini-Hochberg method) (right). (C) Genome browser plots for _Hoxb1_ locus. The
top two rows represent the chromatin accessibility in VEH and in STZ within AHF2. The third track from the top shows the genomic location of the DAR with more accessibility in STZ (red
rectangles, highlighted by yellow box). The second track from the bottom represent the links between peaks and gene (‘Peak2GeneLinks’), calculated by ArchR. Darker lines represent stronger
links. The bottom track shows the gene location and transcriptional direction (red – positive strand; blue – negative strand). EXTENDED DATA FIG. 8 ENHANCED RETINOIC ACID SIGNALING IN
PHARYNGEAL ARCH 2 AND AHF2 IN RESPONSE TO HYPERGLYCEMIA. (A) Box plots of the distribution of ChromVAR deviation score for RAR and RXR transcription factor motifs for each cluster in the
neural crest cell population. PA2 neural crest cells are highlighted in red. The X-axis shows the distribution of the Z-score. (STZ – red; VEH – blue) (n = 6 biological samples. n = 3 VEH
replicates and n = 3 STZ replicates). (B) Box plots of the distribution of ChromVAR deviation score for RAR and RXR transcription factor motifs for each cluster in the mesoderm population.
AHF2 cells are highlighted in red. The X-axis shows the distribution of the Z-score. (STZ – red; VEH – blue) (n = 6 biological samples. n = 3 VEH replicates and n = 3 STZ replicates). In the
box plots, the central line indicates the median, box bounds represent the 25th and 75th percentiles, whiskers extend to values within 1.5 times the interquartile range, and outliers lie
beyond this range. EXTENDED DATA FIG. 9 DISRUPTED RETINOIC ACID SIGNALING IS ASSOCIATED WITH DYSREGULATION OF GENE REGULATORY NETWORKS IN PHARYNGEAL ARCH 2 AND AHF2. (A) Dot plot
demonstrating the distribution of the WGCNA modules per cluster. X axis shows the Z-score differences of WGCNA module score per cluster between STZ and VEH and Y axis shows the statistical
significance of the differences. Modules that are not statistically significant are shown in blue, and those that are statistically significant are shown in green or pink. Red label
highlights selected module used for subsequent analysis. (B) Module scores for a gene module detected in the WGCNA analysis that showed statistically significant variation between VEH and
STZ only in PA2. Linear mixed effects models with mouse id as the random effect was used to test the significance of the mean difference in the module score between VEH and STZ (n = 6
biological samples. n = 3 VEH replicates and n = 3 STZ replicates) (linear mixed-effects model with Benjamini-Hochberg multiple-testing correction). (C) Map of functional protein-protein
interactions (PPI) of genes composing the module described in (B), depicted using STRING. Genes composing a core of the PPI network and being downstream of _Tfap2_ are highlighted in red and
bold. (D) Dot plot demonstrating the distribution of the WGCNA modules per cluster. X axis shows the Z-score differences of WGCNA module score per cluster between STZ and VEH and Y axis
shows the statistical significance of the differences. Modules that are not statistically significant are shown in blue, and those that are statistically significant are shown in green or
pink. Red label highlights selected module used for subsequent analysis. (E) Module scores for a cardiac gene regulatory module detected in the WGCNA analysis that showed statistically
significant variation between VEH and STZ only in AHF2. The same statistical test as (B) was used (n = 6 biological samples. n = 3 VEH replicates and n = 3 STZ replicates) (linear
mixed-effects model with Benjamini-Hochberg multiple-testing correction). (F) Map of PPI of genes composing the module described in (E), depicted using STRING. Genes composing a core of the
PPI network and being critical cardiac TFs or signaling genes are highlighted in red and bold. NC-prog, neural crest cell progenitors; PA2, pharyngeal arch 2; PA3, pharyngeal arch 3; PA4/6,
pharyngeal arch 4/6; SMCs, smooth muscle cells; SMC-Prog, smooth muscle cell progenitors; pSHF1/2, posterior second heart field 1/2; AHF1/2, anterior heart field 1/2; ParaxialMeso1/2,
paraxial mesoderm 1/2. In the box plots, the central line indicates the median, box bounds represent the 25th and 75th percentiles, whiskers extend to values within 1.5 times the
interquartile range, and outliers lie beyond this range. EXTENDED DATA FIG. 10 ANTERIOR EXTENSION OF RETINOIC ACID SIGNALING ACTIVITY IN STZ IN VIVO (COMPLEMENTARY TO FIG. 5E, F). (A) X-gal
staining of RARE-LacZ mouse fetuses at E10.5 from VEH and STZ groups (N = 3 each). Second heart field area and outflow tract area are circled with black lines. Highlighted area in magnified
panels to the right show LacZ positive areas detected by threshold analysis as described in methods. The scale bar represents 1 mm. (B) Measurements of area of second heart field and outflow
tract circled with black line in (A) (N = 3 each). (C) Percentage of LacZ positive area with in the second heart field area and outflow tract area circled with black line in (A) (N = 3
each). Source data SUPPLEMENTARY INFORMATION SUPPLEMENTARY INFORMATION Supplementary Fig. 1, Tables 1–5 and Videos 1–5 and source data for the supplementary figure. REPORTING SUMMARY
SUPPLEMENTARY TABLES SUPPLEMENTARY TABLE 1. CARDIAC PHENOTYPES AT E18.5 FROM VEH- OR STZ-TREATED FEMALES BY MICRO-CT. This table presents the number and types of cardiac phenotypes detected
by micro-CT, including atrial septal defect (ASD), patent foramen ovale (PFO), ventricular septal defect (VSD), atrioventricular septal defect (AVSD) and OFT anomalies, in E18.5 embryos from
VEH- or STZ-treated females after cesarean section. Representative micro-CT images are shown in Extended Data Fig. 1b and Supplementary Video 1a,b. There were significant differences in the
presence of cardiac developmental abnormalities between the VEH and STZ groups by two-sided Fisher’s exact test (_P_ = 0.0004). SUPPLEMENTARY TABLE 2. STATISTICAL RESULTS FOR THE POPULATION
CHANGES IN SCRNA-SEQ AND SCATAC-SEQ DATA FOR FIG. 1G, LEFT (A), FOR FIG. 1G, RIGHT (B), AND FOR FIG. 4B (C). SUPPLEMENTARY TABLE 3. STATISTICAL RESULTS FOR THE CHROMVAR ANALYSIS. This table
presents the statistical results from two-sided Wilcoxon rank-sum test with Benjamini–Hochberg multiple-testing correction to determine the differences between the bias-corrected deviations
for a TF motif between VEH and STZ conditions per each cell type displayed in Extended Data Fig. 9a,b. Gray highlighted cell types show the statistical differences. SUPPLEMENTARY TABLE 4.
PRIMER LISTS. Primers for cloning the candidate distal regulatory regions and deletions discussed in Fig. 5a–d and Extended Data Fig. 7a,b. SUPPLEMENTARY TABLE 5. GUIDE RNA AND HDR TEMPLATE
SEQUENCE USED FOR _ALX3_–CRE TARGET ALLELE GENERATION. SUPPLEMENTARY VIDEO 1A 3D MICRO-CT IMAGES OF E18.5 EMBRYONIC HEARTS FROM VEH AND STZ CONDITIONS. A, 3D micro-CT images of the E18.5
embryonic heart from VEH that is shown in Extended Data Fig. 1b. Pulmonary artery (light green), aorta (light red), right ventricle chamber (dark green) and left ventricle chamber (dark
green) are highlighted. B, 3D micro-CT images of the E18.5 embryonic heart from STZ that is shown in Extended Data Fig. 1b. Pulmonary artery (light green), aorta (light red), right ventricle
chamber (dark green), left ventricle chamber (dark green) and conotruncal ventricular septal defect (purple) are highlighted. SUPPLEMENTARY VIDEO 1B 3D MICRO-CT IMAGES OF E18.5 EMBRYONIC
HEARTS FROM VEH AND STZ CONDITIONS. A, 3D micro-CT images of the E18.5 embryonic heart from VEH that is shown in Extended Data Fig. 1b. Pulmonary artery (light green), aorta (light red),
right ventricle chamber (dark green) and left ventricle chamber (dark green) are highlighted. B, 3D micro-CT images of the E18.5 embryonic heart from STZ that is shown in Extended Data Fig.
1b. Pulmonary artery (light green), aorta (light red), right ventricle chamber (dark green), left ventricle chamber (dark green) and conotruncal ventricular septal defect (purple) are
highlighted. SUPPLEMENTARY VIDEO 2 DISTRIBUTION OF _ALX3_-POSITIVE CELLS IN MEF2C–AHF–CRE:AI6 FETUS AT E10.5. Serial coronal optical sections from ventral to dorsal of whole-mount RNA in
situ hybridization. _Alx3_ (red), ZsGreen (green) and DAPI (blue) are shown. Scale bar, 100 µm. SUPPLEMENTARY VIDEO 3 THE SPATIAL RELATIONSHIP BETWEEN _ALX3_-POSITIVE CELLS AND
_ARMH4_-POSITIVE CELLS. The whole-mount RNA in situ hybridization for _Alx3_ (red), _Armh4_ (green) with DAPI (blue). Scale bar, 300 µm. SUPPLEMENTARY VIDEO 4 THE DISTRIBUTION OF
_ALX3–CRE_:AI6 LINEAGE-TRACED CELLS IN NEONATAL HEARTS. Serial coronal optical sections from dorsal to ventral of the _Alx3_Cre/+:Ai6 mouse neonatal heart shown in Fig. 3g. SUPPLEMENTARY
VIDEO 5A RARE ACTIVITY IS ENHANCED IN THE SECOND HEART FIELD AT E10.5 IN MATERNAL DIABETES. A, The whole-mount RNA in situ hybridization for _Alx3_ (green), LacZ (red) with DAPI (blue) in an
E10.5 RARE–LacZ fetus from VEH-treated female. Scale bar, 300 µm. B, The whole-mount RNA in situ hybridization for _Alx3_ (green), LacZ (red) with DAPI (blue) in an E10.5 RARE–LacZ fetus
from STZ-treated female. Scale bar, 300 µm. SUPPLEMENTARY VIDEO 5B RARE ACTIVITY IS ENHANCED IN THE SECOND HEART FIELD AT E10.5 IN MATERNAL DIABETES. A, The whole-mount RNA in situ
hybridization for _Alx3_ (green), LacZ (red) with DAPI (blue) in an E10.5 RARE–LacZ fetus from VEH-treated female. Scale bar, 300 µm. B, The whole-mount RNA in situ hybridization for _Alx3_
(green), LacZ (red) with DAPI (blue) in an E10.5 RARE–LacZ fetus from STZ-treated female. Scale bar, 300 µm. SOURCE DATA SOURCE DATA FIG. 1 Statistical source data. SOURCE DATA FIG. 4
Statistical source data. SOURCE DATA FIG. 5 Statistical source data. SOURCE DATA EXTENDED DATA FIG. 3 Statistical source data. SOURCE DATA EXTENDED DATA FIG. 10 Statistical source data.
RIGHTS AND PERMISSIONS Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other
rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law. Reprints and
permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Nishino, T., Ranade, S.S., Pelonero, A. _et al._ Single-cell multimodal analyses reveal epigenomic and transcriptomic basis for birth defects
in maternal diabetes. _Nat Cardiovasc Res_ 2, 1190–1203 (2023). https://doi.org/10.1038/s44161-023-00367-y Download citation * Received: 19 August 2022 * Accepted: 19 October 2023 *
Published: 30 November 2023 * Issue Date: December 2023 * DOI: https://doi.org/10.1038/s44161-023-00367-y SHARE THIS ARTICLE Anyone you share the following link with will be able to read
this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative
Trending News
What will the team be for england's next test v sri lanka in may?With the Ashes over, England will not play another Test match until May. Then, they will take on Sri Lanka at Old Traffo...
The truth about the amazons – the real wonder womenThe long-awaited film version of Wonder Woman opens this week, starring Gal Gadot as the indomitable superhero. As Princ...
Page Not Found很抱歉,你所访问的页面已不存在了。 如有疑问,请电邮[email protected] 你仍然可选择浏览首页或以下栏目内容 : 新闻 生活 娱乐 财经 体育 视频 播客 新报业媒体有限公司版权所有(公司登记号:202120748H)...
Help for veterans in suicide crisis | veterans affairsCOMPACT Act (Section 201) Veterans in suicide crisis may be eligible to receive emergency health care services at any VA...
Devoted princes keep diana’s legacy alive 20 years after tragic deathACROSS THE ATLANTIC IT IS ALREADY BEING HAILED AS THE “THERESA EFFECT”... AND IT’S LITTLE WONDER. ON FRIDAY THE WORLD WA...
Latests News
Single-cell multimodal analyses reveal epigenomic and transcriptomic basis for birth defects in maternal diabetesABSTRACT Maternal diabetes mellitus is among the most frequent environmental contributors to congenital birth defects, i...
Spring 2023 tv preview: 14 must-watch showsSpring is not just about warm breezes and fresh blossoms — it’s also about great new TV shows arriving on the networks a...
Gm trails far behind tesla in ev sales — ceo mary barra bet the company that will changeIn this article * GM Follow your favorite stocksCREATE FREE ACCOUNT GM Chair and CEO Mary Barra addresses investors Oct....
Page Not Found很抱歉,你所访问的页面已不存在了。如有疑问,请电邮[email protected]你仍然可选择浏览首页或以下栏目内容 :新闻生活娱乐财经体育视频播客新报业媒体有限公司版权所有(公司登记号:202120748H)...
Labour civil war: john mcdonnell plotted coup against jeremy corbynJust five months after Mr Corbyn officially stepped down as Labour leader, a new book has revealed the conflicts and ten...