Beta-adrenergic agonism protects mitochondrial metabolism in the pancreatectomised rat heart
Beta-adrenergic agonism protects mitochondrial metabolism in the pancreatectomised rat heart"
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT The diabetic heart is characterised by functional, morphological and metabolic alterations predisposing it to contractile failure. Chronic sympathetic activation is a feature of the
pathogenesis of heart failure, however the type 1 diabetic heart shows desensitisation to β-adrenergic stimulation. Here, we sought to understand the impact of repeated
isoprenaline-mediated β-stimulation upon cardiac mitochondrial respiratory capacity and substrate metabolism in the 90% pancreatectomy (Px) rat model of type 1 diabetes. We hypothesised
these hearts would be relatively protected against the metabolic impact of stress-induced cardiomyopathy. We found that individually both Px and isoprenaline suppressed cardiac mitochondrial
respiration, but that this was preserved in Px rats receiving isoprenaline. Px and isoprenaline had contrasting effects on cardiac substrate metabolism, with increased reliance upon cardiac
fatty acid oxidation capacity and altered ketone metabolism in the hearts of Px rats, but enhanced capacity for glucose uptake and metabolism in isoprenaline-treated rats. Moreover, Px rats
were protected against isoprenaline-induced mortality, whilst isoprenaline elevated cGMP and protected myocardial energetic status in Px rat hearts. Our work suggests that adrenergic
stimulation may be protective in the type 1 diabetic heart, and underlines the importance of studying pathological features in combination when modeling complex disease in rodents. SIMILAR
CONTENT BEING VIEWED BY OTHERS SEMAGLUTIDE ADMINISTRATION PROTECTS CARDIOMYOCYTES IN _DB/DB_ MICE VIA ENERGETIC IMPROVEMENT AND MITOCHONDRIAL QUALITY CONTROL Article 24 January 2025 THE
SGLT2 INHIBITOR EMPAGLIFLOZIN IMPROVES CARDIAC ENERGY STATUS VIA MITOCHONDRIAL ATP PRODUCTION IN DIABETIC MICE Article Open access 17 March 2023 CARDIOPROTECTIVE EFFECTS OF EMPAGLIFLOZIN
AFTER ISCHEMIA AND REPERFUSION IN RATS Article Open access 05 May 2021 INTRODUCTION Type 1 diabetes is associated with increased mortality in comparison with the general population, with
cardiovascular diseases being the major cause of death in older patients1. Whilst hypertension and coronary artery disease are prevalent in these patients, the incidence of heart failure
remains high even when these are accounted for. Of note, patients are susceptible to the development of diabetic cardiomyopathy (DbCM), a condition characterised by multiple structural,
functional and metabolic alterations2. There remains a great need to better understand the link between DbCM and heart failure, and in particular how molecular derangements of the diabetic
heart interact with other common components of cardiac pathology3,4. The healthy heart is metabolically flexible, consuming a variety of metabolic substrates to meet ATP requirements
depending on circumstances. Around 60–70% of myocardial ATP demand is met by fatty acid oxidation (FAO), but with an increased reliance on glucose metabolism during the insulin-stimulated,
post-prandial state and in response to stresses such as exercise and hypoxia. Type 1 diabetes greatly increases the reliance of the heart on FAO, reducing the capacity to increase glucose
oxidation in response to metabolic stress5,6. As such, isolated, perfused hearts from type 1 diabetic rats have low glucose oxidation capacities, even when measured in the absence of fatty
acids7. Alongside this impaired flexibility in substrate metabolism, cardiac mitochondrial alterations are typically observed in both animal models of type 1 diabetes and in patients2. For
instance, in rats injected with streptozotocin (STZ) to ablate pancreatic beta-cell function, cardiac mitochondrial respiratory capacity is suppressed8,9,10. This was associated with an
increased ratio of FAO to glucose oxidation capacity when measured in hearts ex vivo8,11,12,13,14, and with increased production of reactive oxygen species (ROS)9,15. Impaired flexibility of
myocardial substrate utilisation and mitochondrial impairments have been extensively associated with impaired contractile function in both patients with advanced heart failure and
experimental rodent models of failure, including those of non-metabolic origin16,17. Chronic sympathetic nervous system activation is a prominent neurohormonal feature of the pathogenesis of
heart failure, and the synthetic β-adrenoreceptor agonist, isoprenaline has been administered experimentally to rodents resulting in a stress-induced cardiomyopathy phenotype18. In rats,
this promotes alterations in cardiac morphology, impaired contractile function and altered metabolism19,20, including a suppression of FAO20. β-adrenergic stimulation might therefore be
expected to further worsen substrate metabolism and energetics in the diabetic heart; however type 1 diabetes is associated with decreased responsiveness to β-stimulation and lower densities
of cardiac β-adrenoreceptors21. In previous work, we superimposed isoprenaline administration onto a surgically induced rat model of type 1 diabetes (90% pancreatectomy; Px) to explore the
impact on cardiac phenotype4. We found that Px alone was associated with myocardial remodelling, but elicited no difference in ejection fraction, whilst isoprenaline decreased ejection
fraction and induced fibrosis4. There was no apparent interaction between the two interventions that worsened cardiac pathology, and whilst first exposure to isoprenaline resulted in 43%
mortality in sham-operated animals, there were no deaths in the Px rats following isoprenaline treatment4. Of note, gene expression analysis indicated downregulation of the β2-adrenoreceptor
(though not the β1-adrenoreceptor) in the hearts of Px rats, suggesting a possible mechanism for this protection4. Here, we sought to understand the cardiac mitochondrial and metabolic
consequences of 90% Px and isoprenaline, both alone and in combination, hypothesising that the hearts of Px rats would be relatively protected against the metabolic consequences of
isoprenaline administration in comparison to those of non-diabetic, sham-operated rats. RESEARCH DESIGN AND METHODS ANIMAL EXPERIMENTATION All work was approved by The Danish Animal
Experiments Inspectorate (license no. 2019-15-0201-01648, 2017-15-0201-01183) and conformed to the European Parliament Directive on the Protection of Animals Used for Scientific Purposes
(2010/63/EU). All experiments were carried out according to the relevant guidelines and regulations. This study is reported in accordance with the ARRIVE guidelines. As reported previously4,
male Sprague–Dawley rats (NTac:SD) (8–10 weeks old, 210–280 g, Taconic Biosciences) were randomised into groups undergoing either removal of 90% of the pancreas (pancreatectomy, Px) or a
sham operation. Following an overnight fast, rats received subcutaneous injections of atropine (0.1 mg/kg), enrofloxacin (50 mg/kg), carprofen (50 mg/kg) and saline (20 ml/kg) prior to
surgery before operation under isoflurane (2–3%) induced anaesthesia. Pancreatectomy (90%) was performed by ligation and then removal of the tail, body and part of the pancreatic head by
gentle abrasion with a dental applicator. The remaining 10% of the pancreas comprised the portion between the duodenal loop and the pancreatic duct. All major blood vessels to the stomach,
spleen and gut were left intact. Sham operation consisted of anaesthesia and ligation of the equivalent portion of the pancreas, before the ligature was removed to leave behind a fully
functional pancreas. Rats received no food from one day before until 1 day after surgery, whereupon they were offered 5 g of chow. From the second day post-surgery normal rodent chow and
water was provided ad libitum. Throughout, rats were housed in conventional cages with a normal 12 h/12 h light/dark photoperiod. Commencing 5 weeks post-operation, pancreatectomised and
sham operated animals were randomised into groups which underwent daily subcutaneous administration of either isoprenaline hydrochloride dissolved in saline (1 mg/kg), or vehicle (saline),
for 10 days. This yielded four study groups: sham-operated rats administered vehicle (Sham/Vehicle), Px rats administered vehicle (Px/Vehicle), sham-operated rates administered isoprenaline
(Sham/Iso), and pancreatectomised rats administered isoprenaline (Px/Iso). Blood glucose and bodyweight were monitored throughout. Tail vein blood samples were collected weekly following a 4
h fast. Samples were collected in heparinised glass capillary tubes, and suspended in glucose/lactate system solution buffer (EKF-diagnostics, Germany). Blood glucose was measured using a
BIOSEN c-Line glucose meter (EKF-diagnostics, Germany). At 10 weeks post-operation, rats were euthanised with rising concentrations of CO2, with death confirmed by cervical dislocation.
Hearts were extracted, and the left ventricle sectioned before being snap frozen for further analysis (RNAseq and mass spectrometry) or transferred fresh into ice-cold biopsy preservation
solution (BIOPS; 2.77 mM CaK2EGTA, 7.23 mM K2EGTA, 6.56 mM MgCl2.6H2O, 20 mM taurine, 15 mM phosphocreatine, 20 mM imidazole, 0.5 mM DTT, 50 mM 4-morpholineethanesulfonic hydrate, and 5.77
mM Na2ATP, pH 7.1.) for immediate analysis of mitochondrial respiratory function using high-resolution respirometry. To ensure statistical independence, only one tissue sample from each rat
was used for each experiment. Each tissue sample, therefore, was considered to be an experimental unit. HIGH-RESOLUTION RESPIROMETRY A section of non-frozen left ventricle was dissected into
bundles of 6–8 fibres, and permeabilised with gentle rocking for 20 min at 4 °C in BIOPS with 50 μg/ml saponin22. Fibre bundles were then washed (3 × 5 min, 4 °C) in respiration medium
(MiR05: 0.5 mM EGTA, 3 mM MgCl2.6H2O, 60 mM K-lactobionate, 20 mM taurine, 10 mM KH2PO4, 20 mM HEPES, 110 mM sucrose, 1 g/l defatted BSA, pH 7.1). Cardiac fibre bundles (2–3 mg) were
immediately added to Oxygraph-O2k chambers (Oroboros Instruments, Innsbruck, Austria) containing 2 ml MiR05 at 37 °C. Respiratory capacities were assessed as described previously22,23.
Malate (2 mM) and octanoyl carnitine (0.2 mM) were initially added to stimulate LEAK respiration (MOctL), followed by 5 mM ADP to stimulate oxidative phosphorylation (OXPHOS) supported by
β-oxidation (MOct_P_). Next, pyruvate (20 mM) was added (MOctPyr_P_) followed by glutamate (10 mM) to assess the capacity for complex I-supported respiration (MOctPyrG_P_), and cytochrome
_c_ (10 µM) to assess outer mitochondrial membrane integrity. Succinate (10 mM) was subsequently added to additionally activate complex II (GMS_P_), followed by 0.5 µM rotenone to inhibit
complex I (S_P_). The ratio of octanoyl carnitine to pyruvate-supported OXPHOS was calculated to indicate relative capacity for fatty acid oxidation (FAO)22. Oxidative coupling efficiency
(OCE) was assessed as (MOct_P_ − MOct_L_)/MOct_P_. MASS SPECTROMETRY Metabolites were extracted from frozen left ventricle using a Bligh–Dyer method as described previously24. Levels of
glycolytic intermediates plus ATP and phosphocreatine (PCr) in the aqueous extract were then assessed using a BEH amide column chromatography method coupled to a Quantiva triple-quadrupole
mass spectrometer (Thermo Scientific). Fractions were reconstituted in an acetonitrile: 10 mM ammonium carbonate solution (7:3 v/v, 100 μl) containing a 10 μM mixture of internal standards
(200 ml; phenylalanine d5, valine d8, leucine d10). The column used was a 1.7 μm BEH amide column (150 × 2.1 mm), coupled to a Vanquish UHPLC + series (Thermo Scientific, UK) LC system and a
TSQ Quantiva Triple Quadrupole Mass Spectrometer (Thermo Scientific). The mobile phase was pumped at 600 μl min-1 with mobile phase A 0.1% ammonium carbonate solution, and mobile phase B
acetonitrile. Mobile phase A was held at 20% for 1.5 min, linearly increased to 60% over the next 2.5 min, held at 60% for 1 min, before being decreased back to the initial conditions (20%
mobile phase A) over 0.1 min. The total run time was 6 min. Nitrogen at 48 mTorr, 420 °C was used as a drying gas for solvent evaporation and the UPLC column was conditioned at 30 °C. The
Quantiva utilised PosNeg switching at voltages of 3.5 and − 2.5 kV for ionisation. GENE EXPRESSION RNAseq was employed with the purpose of analysing the expression of pre-defined target
genes associated with major pathways of substrate metabolism, namely fatty acid uptake/oxidation, glucose uptake/glycolysis, and ketolysis/ketogenesis. RNA was purified from homogenised
tissue using the NucleoSpin RNA Plus kit (Macherey–Nagel GmbH). 500 ng purified RNA from each sample was used to generate a cDNA library using the NEBNext® Ultra™ II Directional RNA Library
Prep Kit from Illumina (New England Biolabs). The cDNA library was then sequenced on a NextSeq 500 using NextSeq 500/550 High Output Kit V2 (Illumina). Sequencing was carried out for 75
cycles and 2 × 8 index cycles. The sequencing data was aligned to the rat genome obtained from the Ensembl database using the Spliced Transcripts Alignment to a Reference (STAR) software.
Gene expression as RPKM is presented relative to the sham/vehicle group. STATISTICS Statistical analysis was performed using a two-way ANOVA, with Isoprenaline treatment and Pancreatectomy
as the variables. Significant differences were further analysed by a Tukey’s post-hoc honest significance difference test. Statistical significance was considered at p < 0.05. ETHICS
APPROVAL AND CONSENT TO PARTICIPATE All work was approved by The Danish Animal Experiments Inspectorate (license no. 2019-15-0201-01648, 2017-15-0201-01183) and conformed to the European
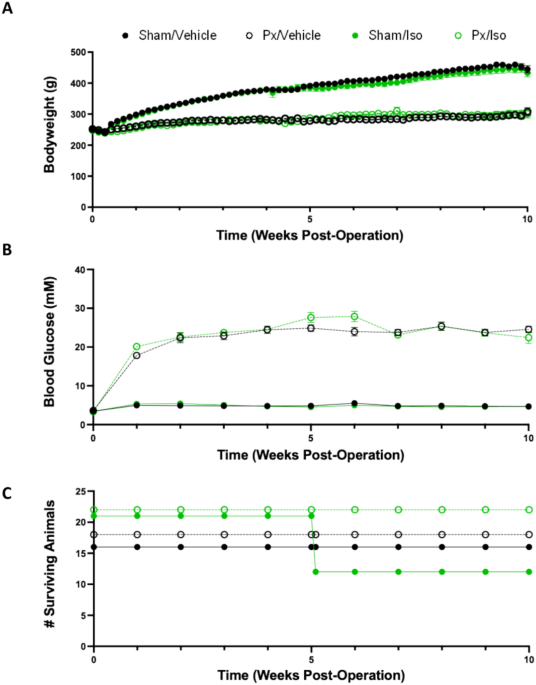
Parliament Directive on the Protection of Animals Used for Scientific Purposes (2010/63/EU). RESULTS BLOOD GLUCOSE, BODY WEIGHT AND MORTALITY Rats which underwent pancreatectomy gained less
weight over the course of the 10 weeks than sham-operated counterparts (Fig. 1A), with final bodyweights for Px rats being 33% lower (p < 0.0001). Px rats reached hyperglycaemia within 1
week post-surgery, with blood glucose concentration reaching a plateau at 2 weeks post-surgery, of 22.4 ± 1.3 and 22.6 ± 1.16 mM for Px/Vehicle and Px/Iso rats, respectively (Fig. 1B).
Sham-operated rats did not exhibit hyperglycaemia, with blood glucose concentrations of 4.9 ± 0.17 mM for Sham/Vehicle and 5.4 ± 0.25 mM for Sham/Iso. Neither blood glucose nor bodyweight
were affected by isoprenaline administration. Pancreatectomised rats were protected against the mortality induced by isoprenaline administration. Following receipt of the first
administration of isoprenaline, 9 of the 21 sham-operated rats in the Sham/Iso group were humanely euthanised or died within the first 24 h following administration. In contrast, all
pancreatectomised rats receiving isoprenaline survived, and as expected there was no mortality associated with vehicle administration (Fig. 1C). CARDIAC MITOCHONDRIAL RESPIRATORY CAPACITY
Both pancreatectomy and isoprenaline administration suppressed aspects of cardiac mitochondrial respiratory capacity, although the nature of this suppression differed between the two
interventions and there was a statistically significant interaction between the two. Pancreatectomy alone, did not suppress LEAK or OXPHOS state respiration supported by octanoyl-carnitine
(MOct_L_ or MOct_P_; Fig. 2A), suggesting a protection of fatty acid oxidation capacity. OXPHOS supported by complex I substrates was suppressed by pancreatectomy, with 28.9% lower
respiration following the addition of pyruvate (MOctPyr_p_, p < 0.01) and 26.9% lower respiration following the addition of glutamate (MOctPyrG_p_, p < 0.01) in comparison with
sham-operated rats. Maximal OXPHOS capacity supported by substrates for complex I and II in combination (GS_P_) was 38.4% lower in the hearts of pancreatectomised rats compared with
sham-operated rats (p < 0.0001), whilst OXPHOS supported by the complex II substrate succinate (S_P_) was 37.7% lower (p < 0.01). Reflecting a relative protection of fatty acid
oxidation in the hearts of pancreatectomised rats, the proportion of complex I activity that could be supported by FAO was 12% greater in the Px/vehicle group relative to sham/vehicle rats
(Fig. 2B; Px effect, _p_ < 0.01). Isoprenaline administration resulted in a statistically significant suppression of mitochondrial respiratory capacity in all states measured (Fig. 2A).
In the hearts of isoprenaline-treated rats, LEAK and OXPHOS respiration in the presence of malate and octanoyl-carnitine (MOct_L_ and MOct_P_) were 15.8% and 25.8% lower (p < 0.05) than
in vehicle-treated rats. Complex I supported respiration rates were 20.1% and 20% lower for MOctPyr_p_ and MOctPyrG_P_, respectively (p < 0.05). Maximal OXPHOS capacity supported by
substrates for complex I and II in combination (GS_P_) was 26.2% lower in the hearts of isoprenaline-treated rats compared with sham-operated rats (p < 0.01), whilst OXPHOS supported by
the complex II substrate succinate (S_P_) was 30.9% lower in isoprenaline administered rats compared with sham-operated controls (p < 0.05). Isoprenaline treatment did not alter the
proportion of complex I respiration that could be supported by FAO (Fig. 2B). There was a statistically significant interaction between the effects of pancreatectomy and isoprenaline for all
respiration rates measured (Px:Iso interaction effect p < 0.001 for all respiration rates, Fig. 2A). As such, the suppression of respiratory capacity seen in pancreatectomised rats
compared with sham-operated controls was not seen in pancreatectomised rats treated with isoprenaline. OXPHOS respiration supported by complex I substrates was 27% higher (MOct_p_, p <
0.05) and 25% higher (MOctPyrG_P_, p < 0.05) in the hearts of Px/Iso rats compared with Px/Veh rats. Maximal OXPHOS capacity supported by substrates for complex I and II in combination
(GS_P_) was 43% higher in the hearts of Px/Iso rats compared with Px/Veh rats (p < 0.01), whilst OXPHOS supported by the complex II substrate succinate (S_P_) was 48% higher (p <
0.05). Neither pancreatectomy nor isoprenaline, alone or in combination, altered mitochondrial OXPHOS coupling efficiency (Fig. 2C). Taken together, these results indicate that whilst
pancreatectomy and isoprenaline independently suppressed components of cardiac mitochondrial respiratory capacity, when applied in combination respiratory capacity was preserved. FATTY ACID
OXIDATION GENES To further investigate the metabolic effects of pancreatectomy and isoprenaline administration, we measured the expression of genes encoding proteins that support myocardial
fatty acid oxidation. Expression of _Cd36_ (encoding the sarcolemmal fatty acid uptake transporter CD36/FAT) was 82% higher in the hearts of pancreatectomised rats compared with
sham-operated controls (Px effect p < 0.0001; Fig. 3A). Similarly, cardiac expression of _Cpt1b_ (encoding carnitine palmitoyl transferase 1, CPT1) was 21% higher (Px effect p <
0.0001) in pancreatectomised rats (Fig. 3b). Expression levels of _Acadvl_, _Hadh_, _Acat_ and _Acadm_ (encoding β-oxidation enzymes) were respectively 35%, 33%, 51% and 21% higher (Px
effect p < 0.0001) in the hearts of pancreatectomised rats than in those of sham-operated rats (Fig. 3C–F). There was no significant effect of isoprenaline administration on the
expression of any of these genes. In conjunction with our respirometry results, these findings therefore suggest a relative preservation of FAO capacity in the hearts of pancreatectomised
rats. GLUCOSE METABOLISM To further probe the effects of pancreatectomy and isoprenaline administration on cardiac substrate metabolism we measured the levels of glycolytic intermediates,
and the expression of genes associated with glucose uptake and metabolism (Fig. 4). Expression of _Glut1_ (encoding the constitutive glucose uptake transporter GLUT1) was suppressed by
pancreatectomy (Px effect p < 0.001) but enhanced by isoprenaline (Iso effect p < 0.001), such that expression levels were 58% lower in pancreatectomised rats compared with
sham-operated controls, but 41% higher in isoprenaline-treated rats compared with vehicle-treatment. Myocardial concentrations of early glycolytic intermediates were elevated in
pancreatectomised rats compared with sham-operated controls (Px effect p < 0.01), with 65% higher levels of glucose-6-phosphate and 38% higher levels of fructose bisphosphate detected in
these hearts. There was no significant effect of isoprenaline on these intermediates. Myocardial expression of _Aldoc_ (encoding aldolase C) was significantly enhanced by isoprenaline
administration (Iso effect p < 0.001) with expression following isoprenaline treatment being 43% higher in the hearts of sham-operated animals and 100% higher in pancreatectomised rats.
Furthermore, isoprenaline treatment significantly enhanced myocardial concentrations of glycolytic intermediates downstream of aldolase (Iso effect p < 0.01), with levels of
dihydroxyacetone, glyceraldehyde-3-phosphate and 2/3-phosphoglycerate being 42%, 38% and 54% higher, respectively, in the hearts of isoprenaline-treated sham-operated rats, compared with
vehicle-treated controls. Similarly, concentrations of the same three metabolites were respectively 35%, 42% and 40% higher in pancreatectomised rats following isoprenaline treatment,
compared with vehicle-treated counterparts. Isoprenaline administration also resulted in lower expression of genes encoding two pyruvate dehydrogenase kinase isoforms (_Pdk1_ and _Pdk2_; Iso
effect _p_ < 0.01 and _p_ < 0.05, respectively), potentially supporting greater rates of pyruvate flux through pyruvate dehydrogenase. Myocardial _Pdk1_ expression was respectively
10% and 18% lower following isoprenaline administration in sham-operated and pancreatectomised rats respectively, whilst _Pdk2_ expression was 6% and 14% lower in the same hearts. To
investigate metabolite levels in pathways into which glycolytic intermediates might be diverted, levels of pentose phosphate pathway metabolites, and of lactate, were also measured by MS.
6-phosphoglycerate levels were 31.4% higher in Px/Vehicle rats vs Sham/Vehicle controls, and 4.03-fold greater in LV from Px/Iso rats than in Sham/Iso LV (Supplementary Fig. 1A, Px effect,
_p_ < 0.05). There was no effect of either treatment upon ribulose-5-phosphate levels (Supplementary Fig. 1B), but ribose-5-phosphate levels were 40.5% higher in Sham/Iso rats vs
Sham/Vehicle controls, and 63.1% higher in LV from Px/Iso rats than Px/Vehicle LV (Supplementary Fig. 1C, Iso effect, _p_ < 0.05). Lactate levels were unchanged between the groups
(Supplementary Fig. 1D). These findings collectively point towards a greater capacity for glycolysis, and possible relative protection of pyruvate oxidation in the hearts of isoprenaline
treated rats. KETONE BODY METABOLISM Next, we measured levels of the ketone body, β-hydroxybutyrate (β-OHB), and the expression of genes encoding enzymes that support myocardial ketone
metabolism. Pancreatectomy resulted in elevated myocardial β-hydroxybutyrate (β-OHB) levels (Fig. 5A; Px effect _p_ < 0.01), which were 41% greater in the hearts of pancreatectomised rats
administered vehicle and 19% greater in those treated with isoprenaline, compared with the respective sham-operated controls. Expression of both _Hmgcl_ (encoding HMG-CoA Lyase) and
_Hmgcs2_ (encoding HMG-CoA Synthase 2) were increased in the hearts of pancreatectomised rats (Fig. 5B,C; Px effect of _p_ < 0.0001 for both targets). As such, _Hmgcl_ expression was
1.4-fold higher in the hearts of pancreatectomised rats compared with sham-operated controls, whilst expression of _Hmgcs2_ was 14.2-fold higher in the same hearts. In contrast, expression
of _Bdh1_ (encoding β-hydroxybutyrate dehydrogenase) and _Oxct1_ (encoding succinyl-CoA:3-ketoacid CoA transferase) were suppressed in the hearts of pancreatectomised rats compared with
sham-operated rats (Fig. 5D,E; Px effect of _p_ < 0.0001 for both targets). Expression of _Bdh1_ was 74% lower in the hearts of vehicle-administered, pancreatectomised rats and 83% lower
in those treated with isoprenaline, whilst _Oxct1_ expression was respectively 58% and 60% lower in the same hearts. MYOCARDIAL HIGH-ENERGY PHOSPHATE AND CYCLIC NUCLEOTIDE LEVELS We next
considered phosphocreatine (PCr) and ATP levels in the hearts of pancreatectomised and isoprenaline treated rats. Px/Vehicle rats exhibited 35% lower myocardial PCr levels than Sham/Vehicle
rats (Fig. 6A), though no difference in ATP levels (Fig. 6B). PCr levels in Px/Iso rat hearts were not lower than in Sham/Vehicle rats however (Fig. 6A; Iso effect, _p_ < 0.01), and had
2.4-fold higher levels of ATP (Fig. 6B, Iso effect, _p_ < 0.05. Isoprenaline administration thereby appeared to increase high energy phosphate availability; a finding that aligned with
the protection of mitochondrial respiratory capacity in the hearts of these rats. Following isoprenaline administration, levels of AMP and ADP levels were 15.0% and 31.0% higher respectively
in Sham/Iso rat LV vs Sham/Vehicle (Supplementary Fig. 2A,B, Iso effects of _p_ < 0.05 and _p_ < 0.01 respectively). Creatine levels, meanwhile, were 30.9% greater in Px/Vehicle LV
than in Sham/Vehicle LV (Supplementary Fig. 2C, Px effect, _p_ < 0.01). Finally, to understand the mechanistic basis for our findings and considering our previous data showing that
pancreatectomy altered the myocardial expression of genes encoding mediators of adrenergic signalling and cyclic nucleotide metabolism4, we sought to understand the impact of pancreatectomy
and isoprenaline on myocardial levels of cAMP and cGMP. We found that neither pancreatectomy nor isoprenaline had a significant effect on myocardial cAMP levels (Fig. 6C), however
isoprenaline increased cGMP levels in rat heart (Fig. 6D; Iso effect of p < 0.01). DISCUSSION In this study, we sought to understand the metabolic consequences of β-adrenergic
stimulation, via repeated isoprenaline administration, on cardiac substrate metabolism in the 90% pancreatectomy rat model of type 1 diabetes. We found that pancreatectomy resulted in a
suppression of cardiac mitochondrial respiratory capacity, but a relative preservation of fatty acid oxidation capacity and altered expression of genes associated with ketone body
metabolism. Isoprenaline administration also suppressed mitochondrial respiratory capacity in the hearts of sham-operated rats, but in contrast preserved oxidative phosphorylation capacity
in the hearts of pancreatectomised rats. Moreover, isoprenaline altered the expression of genes associated with glucose metabolism alongside levels of myocardial glycolytic intermediates,
and increased myocardial cGMP levels. A strength of this study was the use of the 90% pancreatectomy rat model, which recapitulates many of the features of type 1 diabetes including
hyperglycaemia, hypoinsulinaemia and poor glucose tolerance following a reduction in β-cell mass. This model of type 1 diabetes holds advantages over other models for the study of diabetic
cardiomyopathy, in that the direct toxic effects associated with streptozotocin administration on tissues including the heart2,25,26,27 are avoided, along with the confounding effects of
obesity and/or altered leptin signalling seen in models such as the ZDF rat. Limitations of this surgical model, however, include the abrogation of pancreatic endocrine and exocrine
functions beyond effects on insulin secretion. In particular, the impact of pancreatectomy on digestive enzyme secretion could include poor nutrient absorption, weight loss and malnutrition,
whilst loss of endocrine glucagon signalling, including cardioprotective effects28 and effects on tissue metabolism29, may also be a feature of this model. A further strength of this study
was the detailed metabolic phenotyping we carried out at the tissue level, including high-resolution respirometry for the analysis of mitochondrial respiratory capacity under different
substrate-led pathways, alongside targeted metabolite analysis via mass spectrometry and metabolic gene expression. Whilst respirometry, carried out ex vivo in the presence of saturating
concentrations of substrates and oxygen, reflects capacity rather than metabolic flux in vivo, there was agreement between these measurements and those derived from other techniques. For
instance, the relative preservation of mitochondrial fatty acid oxidation capacity seen in permeabilised cardiac fibres from pancreatectomised rats was reflected in the elevated expression
of genes encoding enzymes of β-oxidation in these same hearts. Respirometry in permeabilised fibres does not however allow for the measurement of glycolytic flux, which we have instead
investigated by measuring concentrations of glycolytic intermediates, and the expression of glycolytic enzymes by LC–MS and RNAseq. We do, however, acknowledge that a limitation of our study
is the lack of true measurements of metabolic flux, which could for example be made using stable isotopes, and future studies might investigate some of the pathways we highlight here in a
more targeted manner using this approach. In previous work we reported that isoprenaline did not exacerbate the pathological remodeling associated with type 1 diabetes in the hearts of 90%
pancreatectomised rats. In line with this, pancreatectomised rats were protected against isoprenaline-induced mortality in comparison with their sham-operated counterparts4. As seen in this
study and previous work18,19,20, isoprenaline administration has been recorded to induce mortality, albeit often at higher doses. The mechanism for this is not known, however the findings of
this study may shed some light on the mechanism, since pancreatectomy was found to abrogate the effect. This protection may have arisen as a consequence of the downregulation of ADRB2
(encoding the β2-adrenoreceptor) in these rats4, decreasing myocardial sensitivity to adrenergic stimulation, in line with previous findings in the diabetic heart21. Indeed, isoprenaline was
found to induce a weaker inotropic response in type 1 diabetic rat hearts than in non-diabetic hearts, both in vivo and ex vivo30,31. Isoprenaline stimulation of the heart has been reported
to be augmented by enhanced pancreatic secretion of glucagon32, itself an inotropic agent, and as such abrogated glucagon production may be another mechanism of protection in
pancreatectomised rats. The metabolic consequences of the two interventions employed here differed greatly and did not appear to be exacerbated when isoprenaline administration was
superimposed upon pancreatectomised rats. Pancreatectomy was associated with a relative protection of mitochondrial fatty acid oxidation capacity, in the face of overall suppression of
mitochondrial respiratory capacity, and enhanced expression of genes supporting fatty acid oxidation. These findings correspond with reports in other rodent models of diabetes33,34, in
particular STZ-induced models of type 1 diabetes35, with the mechanism likely to involve stimulation of the fatty acid-activated transcription factor, peroxisome proliferator-activated
receptor α (PPARα)14. In addition, pancreatectomy resulted in the altered expression of genes involved in ketone body metabolism, including a marked upregulation of _Hmgcs2_, which encodes
the canonically-ketogenic enzyme HMGCS2, and downregulation of _Oxct_, which encodes the ketolytic enzyme SCOT. Increased expression of HMGCS2 was previously reported in the hearts of
STZ-induced type 1 diabetic rat hearts36, and this may have consequences for the susceptibility of the diabetic heart to ischaemia–reperfusion injury37. In contrast with the effects of
pancreatectomy, repeated isoprenaline administration did not specifically alter the myocardial expression of genes associated with fatty acid oxidation or ketone body metabolism; however it
did result in an overall suppression of mitochondrial respiratory capacity, in conjunction with our previous report of suppression of left ventricular ejection fraction in this model4. This
finding corresponds to a previous report describing the suppression of both fatty acid oxidation and markers of mitochondrial content to a similar degree in the rat heart following chronic
treatment with isoprenaline via osmotic minipumps20. Similar observations of suppressed oxidative metabolism have been made in the chronically-infarcted rat heart, which further exhibited
lower rates of glycolysis38,39. In the present study, we did not find evidence of an overall suppression of glucose metabolism following isoprenaline administration, but instead found
increased expression of _Aldoc_ (encoding aldolase C), _Pdk1_ and _Pdk2_, alongside enhanced levels of glycolytic intermediates downstream of aldolase. We did not see differences in left
ventricular lactate concentrations, however lactate can be rapidly exported from myocardium, and we did not measure lactate efflux. Isoprenaline did increase cardiac levels of high energy
phosphates. The mechanism behind this isoprenaline-mediated increase in ATP is unknown, however levels of AMP and ADP, alongside the later metabolites in the pentose phosphate pathway, were
also raised, hinting that increased levels of purine synthesis may play a role. These findings may collectively be suggestive of altered glycolysis in the face of suppressed oxidative
metabolism, but this would need to be confirmed by direct measurements of glycolytic flux. Of note, both pancreatectomy and isoprenaline administration independently suppressed overall
mitochondrial respiratory capacity in the rat heart, but when isoprenaline was superimposed upon pancreatectomy, mitochondrial respiratory capacity was preserved. The mechanistic basis for
this finding may again lie in the contrasting effects of the two interventions on adrenergic signaling, with pancreatectomy resulting in desensitisation to adrenergic overload following
sustained isoprenaline administration, but isoprenaline stimulation offsetting the accompanying metabolic suppression of type 1 diabetes to some degree through enhanced myocardial cGMP
levels. Indeed, cGMP is a mediator of mitochondrial biogenesis in mammalian cells and tissues, acting at least in part through PPARγ co-activator 1α (PGC1α)40. Augmentation of cGMP signaling
has been proposed as a therapeutic strategy for heart failure with preserved ejection fraction (HFpEF)41, and this might be achieved through phosphodiesterase (PDE) inhibition42,43 or
administration of nitrate/NO-mimetics44,45. Moreover, increased myocardial cGMP signalling has been proposed as a possible mechanism underlying the therapeutic benefits of SGLT2
inhibition46, and this was associated with enhanced fatty acid oxidation and overall ATP production in the hearts of type 2 diabetic (_db/db_) mice47. Our findings shed new light on the
interaction between diabetes-associated myocardial alterations and other factors driving cardiac pathology, underlining the importance of investigating such features in combination. Of note,
an investigation that superimposed pressure overload (via transverse aortic constriction, TAC) onto _db/db_ mice revealed that the diabetic mice were relatively protected against the
functional consequences of TAC in comparison with non-diabetic counterparts3. More strikingly, whilst TAC resulted in energetic and metabolic perturbations in the hearts of non-diabetic
mice, it led to the restoration of cardiac glucose uptake and high-energy phosphate levels in the hearts of _db/db_ mice3, inviting the question of whether two wrongs can make a right when
it comes to metabolic arithmetic48. Our work has implications for the future use of the model we have deployed here and for possible therapeutic avenues targeting metabolism in the diabetic
heart. Previously we sought to investigate the potential of combining pancreatectomy with isoprenaline to establish a novel “dual-hit” model of diabetic cardiomyopathy4. Taken together, the
findings we described previously and here reveal differing structural, functional and metabolic effects associated with the two stressors, and no exacerbation of the pathology with the
combined insult. Moreover, the protection of mitochondrial respiratory capacity in the hearts of isoprenaline-treated pancreatectomised rats, alongside the prevention of isoprenaline-induced
mortality following pancreatectomy, have given us cause to re-evaluate this model. Conversely, our work lends support to therapeutic strategies aiming to allay the metabolic/mitochondrial
derangements in the diabetic heart through the elevation of cGMP, via routes that may include NO/nitrate supplementation or GLP-1/glucagon signaling. CONCLUSION The induction of type-1
diabetes via pancreatectomy and chronic β-adrenergic stimulation through isoprenaline have contrasting effects on myocardial substrate metabolism. Moreover, whilst both stressors suppress at
least some aspects of cardiac mitochondrial respiratory capacity, when administered in combination respiratory capacity is protected. Our work underlines the importance of studying such
stressors in combination when modeling cardiac pathology in rodents, and in particular when considering downstream metabolic consequences. DATA AVAILABILITY Data from this study is available
upon publication at the University of Cambridge Online Data Repository using the following link: https://doi.org/10.17863/CAM.107569. The corresponding author may be contacted to request
data from the study. REFERENCES * Lind, M. _et al._ Glycemic control and excess mortality in type 1 diabetes. _N. Engl. J. Med._ 371, 1972–1982 (2014). Article PubMed Google Scholar *
Bugger, H. & Abel, E. D. Molecular mechanisms of diabetic cardiomyopathy. _Diabetologia_ 57, 660–671 (2014). Article CAS PubMed PubMed Central Google Scholar * Abdurrachim, D. _et
al._ Diabetic db/db mice do not develop heart failure upon pressure overload: A longitudinal in vivo PET, MRI, and MRS study on cardiac metabolic, structural, and functional adaptations.
_Cardiovasc. Res._ 113, 1148–1160 (2017). Article CAS PubMed Google Scholar * Thisted, L. _et al._ Rat pancreatectomy combined with isoprenaline or uninephrectomy as models of diabetic
cardiomyopathy or nephropathy. _Sci. Rep._ 10, 1–15 (2020). Article Google Scholar * Bayeva, M., Sawicki, K. T. & Ardehali, H. Taking diabetes to heart—Deregulation of myocardial
lipid. _J. Am. Heart Assoc._ 2, e000433 (2013). Article PubMed PubMed Central Google Scholar * Stanley, W. C., Lopaschuk, G. D. & McCormack, J. G. Regulation of energy substrate
metabolism in the diabetic heart. _Cardiovasc. Res._ 34, 25–33 (1997). Article CAS PubMed Google Scholar * Wall, S. R. & Lopaschuk, G. D. Glucose oxidation rates in fatty
acid-perfused isolated working hearts from diabetic rats. _Biochim. Biophys. Acta_ 1006, 97–103 (1989). Article CAS PubMed Google Scholar * Flarsheim, C. E., Grupp, I. L. & Matlib,
M. A. Mitochondrial dysfunction accompanies diastolic dysfunction in diabetic rat heart. _Am. J. Physiol. Heart Circ. Physiol._ 271, H192–H202 (1996). Article CAS Google Scholar * Lashin,
O. M., Szweda, P. A., Szweda, L. I. & Romani, A. M. P. Decreased complex II respiration and HNE-modified SDH subunit in diabetic heart. _Free Radic. Biol. Med._ 40, 886–896 (2006).
Article CAS PubMed Google Scholar * Lashin, O. & Romani, A. Hyperglycemia does not alter state 3 respiration in cardiac mitochondria from type-I diabetic rats. _Mol. Cell. Biochem._
267, 31–37 (2004). Article CAS PubMed Google Scholar * How, O. J. _et al._ Rosiglitazone treatment improves cardiac efficiency in hearts from diabetic mice. _Arch. Physiol. Biochem._
113, 211–220 (2007). Article CAS PubMed Google Scholar * Chatham, J. C. & Forder, J. R. Relationship between cardiac function and substrate oxidation in hearts of diabetic rats. _Am.
J. Physiol._ 273, H52–H58 (1997). CAS PubMed Google Scholar * Depre, C. _et al._ Streptozotocin-induced changes in cardiac gene expression in the absence of severe contractile
dysfunction. _J. Mol. Cell. Cardiol._ 32, 985–996 (2000). Article CAS PubMed Google Scholar * Finck, B. N. _et al._ The cardiac phenotype induced by PPARα overexpression mimics that
caused by diabetes mellitus. _J. Clin. Investig._ 109, 121–130 (2002). Article CAS PubMed PubMed Central Google Scholar * Yao, D. & Brownlee, M. Hyperglycemia-induced reactive
oxygen species increase expression of the receptor for advanced glycation end products (RAGE) and RAGE ligands. _Diabetes_ 59, 249–255 (2010). Article CAS PubMed Google Scholar * Murray,
A. J., Edwards, L. M. & Clarke, K. Mitochondria and heart failure. _Curr. Opin. Clin. Nutr. Metab. Care_ 10, 704–711 (2007). Article CAS PubMed Google Scholar * Glatz, J. F. C. _et
al._ Re-balancing cellular energy substrate metabolism to mend the failing heart. _Biochim. Biophys. Acta Mol. Basis Dis._ 1866, 165579 (2020). Article CAS PubMed Google Scholar * Chang,
S. C., Ren, S., Rau, C. D. & Wang, J. J. Isoproterenol-induced heart failure mouse model using osmotic pump implantation. _Methods Mol. Biol._ 1816, 207 (2018). Article CAS PubMed
PubMed Central Google Scholar * Desrois, M. _et al._ Effect of isoproterenol on myocardial perfusion, function, energy metabolism and nitric oxide pathway in the rat heart—A longitudinal
MR study. _NMR Biomed._ 27, 529–538 (2014). Article CAS PubMed Google Scholar * Heather, L. C. _et al._ Isoproterenol induces in vivo functional and metabolic abnormalities; similar to
those found in the infarcted rat heart. _J. Physiol. Pharmacol._ 60, 31–39 (2009). CAS PubMed Google Scholar * Dinçer, Ü. D. _et al._ The effect of diabetes on expression of β1-, β2-, and
β3-adrenoreceptors in rat hearts. _Diabetes_ 50, 455–461 (2001). Article PubMed Google Scholar * Horscroft, J. A. _et al._ Inorganic nitrate, hypoxia, and the regulation of cardiac
mitochondrial respiration—Probing the role of PPARα. _FASEB J._ 33, 7563–7577 (2019). Article CAS PubMed PubMed Central Google Scholar * Horscroft, J. A., Burgess, S. L., Hu, Y. &
Murray, A. J. Altered oxygen utilisation in rat left ventricle and soleus after 14 days, but not 2 days, of environmental hypoxia. _PLoS ONE_ 10, e0138564 (2015). Article PubMed PubMed
Central Google Scholar * Le Belle, J. E., Harris, N. G., Williams, S. R. & Bhakoo, K. K. A comparison of cell and tissue extraction techniques using high-resolution 1H-NMR
spectroscopy. _NMR Biomed._ 15, 37–44 (2002). Article PubMed Google Scholar * Bolzán, A. D. & Bianchi, M. S. Genotoxicity of streptozotocin. _Mutat. Res. Mutat. Res._ 512, 121–134
(2002). Article PubMed Google Scholar * Camacho, P., Fan, H., Liu, Z. & He, J. Q. Small mammalian animal models of heart disease. _Am. J. Cardiovasc. Dis._ 6, 70–80 (2016). CAS
PubMed PubMed Central Google Scholar * Wold, L. E. & Ren, J. Streptozotocin directly impairs cardiac contractile function in isolated ventricular myocytes via a p38 map
kinase-dependent oxidative stress mechanism. _Biochem. Biophys. Res. Commun._ 318, 1066–1071 (2004). Article CAS PubMed Google Scholar * Lindsay, R. T., Ambery, P., Jermutus, L. &
Murray, A. J. Glucagon and exenatide improve contractile recovery following ischaemia/reperfusion in the isolated perfused rat heart. _Physiol. Rep._ 11, e15597 (2023). Article CAS PubMed
PubMed Central Google Scholar * Boland, M. L. _et al._ Resolution of NASH and hepatic fibrosis by the GLP-1R and GCGR dual-agonist cotadutide via modulating mitochondrial function and
lipogenesis. _Nat. Metab._ 2, 413–431 (2020). Article CAS PubMed PubMed Central Google Scholar * Goetzsche, O. L. E. The adrenergic β-receptor adenylate cyclase system in heart and
lymphocytes from streptozotocin-diabetic rats: In vivo and in vitro evidence for a desensitized myocardial β-receptor. _Diabetes_ 32, 1110–1116 (1983). Article CAS Google Scholar * Roth,
D. A., White, C. D., Hamilton, C. D., Hall, J. L. & Stanley, W. C. Adrenergic desensitization in left ventricle from streptozotocin diabetic swine. _J. Mol. Cell. Cardiol._ 27, 2315–2325
(1995). Article CAS PubMed Google Scholar * Almira, E. C. & Misbin, R. I. Effects of insulin and streptozotocin-diabetes on isoproterenol-stimulated cyclic AMP production in
myocytes isolated from rat heart. _Metabolism_ 38, 102–103 (1989). Article CAS PubMed Google Scholar * Makrecka-Kuka, M. _et al._ Altered mitochondrial metabolism in the
insulin-resistant heart. _Acta Physiol._ 228, e13430 (2020). Article CAS Google Scholar * Heather, L. C. _et al._ Guidelines in cardiovascular research: Guidelines on models of diabetic
heart disease. _Am. J. Physiol. Heart Circ. Physiol._ 323, H176 (2022). Article CAS PubMed PubMed Central Google Scholar * Christe, M. E. & Rodgers, R. L. Cardiac glucose and fatty
acid oxidation in the streptozotocin-induced diabetic spontaneously hypertensive rat. _Hypertension_ 25, 235–241 (1995). Article CAS PubMed Google Scholar * Cook, G. A., Lavrentyev, E.
N., Pham, K. & Park, E. A. Streptozotocin diabetes increases mRNA expression of ketogenic enzymes in the rat heart. _Biochim. Biophys. Acta Gen. Subj._ 1861, 307–312 (2017). Article CAS
PubMed Google Scholar * Lindsay, R. T. _et al._ B-hydroxybutyrate accumulates in the rat heart during low-flow ischaemia with implications for functional recovery. _Elife_ 10, 1–16
(2021). Article Google Scholar * Murray, A. J. _et al._ Insulin resistance, abnormal energy metabolism and increased ischemic damage in the chronically infarcted rat heart. _Cardiovasc.
Res._ 71, 149–157 (2006). Article ADS CAS PubMed Google Scholar * Heather, L. C. _et al._ Fatty acid transporter levels and palmitate oxidation rate correlate with ejection fraction in
the infarcted rat heart. _Cardiovasc. Res._ 72, 430–437 (2006). Article CAS PubMed Google Scholar * Nisoli, E. _et al._ Mitochondrial biogenesis by NO yields functionally active
mitochondria in mammals. _Proc. Natl. Acad. Sci. U.S.A._ 101, 16507–16512 (2004). Article ADS CAS PubMed PubMed Central Google Scholar * Greene, S. J. _et al._ The cGMP signaling
pathway as a therapeutic target in heart failure with preserved ejection fraction. _J. Am. Heart Assoc._ 2, 1 (2013). Article Google Scholar * Kukreja, R. C., Salloum, F. N. & Das, A.
Role of cGMP signaling and phosphodiesterase-5 inhibitors in cardioprotection. _J. Am. Coll. Cardiol._ 59, 1921 (2012). Article CAS PubMed PubMed Central Google Scholar * Baliga, R. S.
_et al._ Phosphodiesterase 2 inhibition preferentially promotes NO/guanylyl cyclase/cGMP signaling to reverse the development of heart failure. _Proc. Natl. Acad. Sci. U.S.A._ 115,
E7428–E7437 (2018). Article CAS PubMed PubMed Central Google Scholar * Kojda, G. _et al._ Low increase in cGMP induced by organic nitrates and nitrovasodilators improves contractile
response of rat ventricular myocytes. _Circ. Res._ 78, 91–101 (1996). Article CAS PubMed Google Scholar * Boerrigter, G., Lapp, H. & Burnett, J. C. Modulation of cGMP in heart
failure: A new therapeutic paradigm. _Handb. Exp. Pharmacol._ 191, 485–506 (2009). Article CAS Google Scholar * Xue, M. _et al._ Empagliflozin prevents cardiomyopathy via sGC-cGMP-PKG
pathway in type 2 diabetes mice. _Clin. Sci._ 133, 1705–1720 (2019). Article CAS Google Scholar * Verma, S. _et al._ Empagliflozin increases cardiac energy production in diabetes: Novel
translational insights into the heart failure benefits of SGLT2 inhibitors. _JACC Basic Transl. Sci._ 3, 575–587 (2018). Article PubMed PubMed Central Google Scholar * Lygate, C. A.
Metabolic arithmetic: Do two wrongs make a right? _Cardiovasc. Res._ 113, 1093–1095 (2017). Article CAS PubMed Google Scholar Download references ACKNOWLEDGEMENTS The authors thank Alice
Sowton for her contribution to data analysis. FUNDING The study was funded by Gubra AS. RTL was supported by a British Heart Foundation Grant FS/14/59/31282. AJM was supported by Research
Councils UK Fellowship EP/E500552/1. AUTHOR INFORMATION Author notes * Nora E. Zois Present address: Ascendis Pharma A/S, Hellerup, Denmark * James A. West Present address: AstraZeneca,
Cambridge, UK * Keld Fosgerau Present address: Pephexia Therapeutics ApS, Copenhagen, Denmark * Julian L. Griffin Present address: The Rowett Institute, University of Aberdeen, Aberdeen, UK
* Lisbeth N. Fink Present address: Ferring Pharmaceuticals A/S, Kastrup, Denmark AUTHORS AND AFFILIATIONS * Department of Physiology, Development and Neuroscience, University of Cambridge,
Cambridge, UK Ross T. Lindsay & Andrew J. Murray * Gubra A/S, Hørsholm Kongevej 11B, 2970, Hørsholm, Denmark Ross T. Lindsay, Louise Thisted, Nora E. Zois, Sebastian T. Thrane, Keld
Fosgerau & Lisbeth N. Fink * Department of Biochemistry and Systems Biology Centre, University of Cambridge, Cambridge, UK Ross T. Lindsay, James A. West & Julian L. Griffin Authors
* Ross T. Lindsay View author publications You can also search for this author inPubMed Google Scholar * Louise Thisted View author publications You can also search for this author inPubMed
Google Scholar * Nora E. Zois View author publications You can also search for this author inPubMed Google Scholar * Sebastian T. Thrane View author publications You can also search for this
author inPubMed Google Scholar * James A. West View author publications You can also search for this author inPubMed Google Scholar * Keld Fosgerau View author publications You can also
search for this author inPubMed Google Scholar * Julian L. Griffin View author publications You can also search for this author inPubMed Google Scholar * Lisbeth N. Fink View author
publications You can also search for this author inPubMed Google Scholar * Andrew J. Murray View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS
RTL, LT, NEZ, KF, LF, and AJM designed the study. RTL, LT, NEZ executed the study. RTL, LT, NEZ, STT, JAW, JLG and AJM analysed and interpreted the data from the study. RTL, AJM drafted the
manuscript. All authors reviewed and revised the manuscript. All authors have approved the manuscript for submission. CORRESPONDING AUTHOR Correspondence to Ross T. Lindsay. ETHICS
DECLARATIONS COMPETING INTERESTS LT, NEZ, STT, KF and LF were employees of Gubra ApS at the time of work for this manuscript being undertaken. RTL undertook a temporary, unpaid secondment
with Gubra ApS at this time. RTL is currently an employee and shareholder of Rejuveron AG and declares no other potential or perceived conflicts of interest. NEZ is currently an employee and
shareholder of Ascendis Pharma A/S and declares no potential or perceived conflicts of interest. LNF is currently an employee of Ferring Pharmaceuticals A/S and declares no further
potential or perceived conflicts of interest. KF is currently an employee and shareholder of Pephexia Therapeutics and declares no further potential or perceived conflicts of interest. JAW
is currently an employee of AstraZeneca, and declares no further potential or perceived conflicts of interest. JLG and AJM declare that no conflicts of interest exist. ADDITIONAL INFORMATION
PUBLISHER'S NOTE Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. SUPPLEMENTARY INFORMATION SUPPLEMENTARY FIGURES.
RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and
reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes
were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material.
If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to
obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS
ARTICLE Lindsay, R.T., Thisted, L., Zois, N.E. _et al._ Beta-adrenergic agonism protects mitochondrial metabolism in the pancreatectomised rat heart. _Sci Rep_ 14, 19383 (2024).
https://doi.org/10.1038/s41598-024-70335-4 Download citation * Received: 08 March 2024 * Accepted: 14 August 2024 * Published: 21 August 2024 * DOI:
https://doi.org/10.1038/s41598-024-70335-4 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not
currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative KEYWORDS * Mitochondria * Pancreatectomy * Adrenergic * Heart *
Diabetes * Cardiomyopathy
Trending News
20 New Novels for Spring_WHEN A STRANGER COMES TO TOWN_ BY THE MYSTERY WRITERS OF AMERICA It's said that all great stories are either about...
Diego simeone praises thomas tuchel ahead of atletico’s clash against chelseaAtletico Madrid boss Diego Simeone has heaped praise on Chelsea manager Thomas Tuchel ahead of Tuesday’s Champions Leagu...
First convicts to fight in ukraine war freed by putin ally, urged not to kill or rape againEXPLORE MORE Yevgeny Prigozhin, the founder of Russia’s most powerful mercenary group, bade farewell on Thursday to form...
Best fall fashion trends for women over 50 in 2024 | members onlyMemorial Day Sale! Join AARP for just $11 per year with a 5-year membership Join now and get a FREE gift. Expires 6/4 G...
2 of 3 suspects bond out in jennifer farber dulos murder caseFor Fotis Dulos, who has been charged with murder in the killing of his estranged wife, he posted $420,150, or 7 percent...
Latests News
Beta-adrenergic agonism protects mitochondrial metabolism in the pancreatectomised rat heartABSTRACT The diabetic heart is characterised by functional, morphological and metabolic alterations predisposing it to c...
Page Not FoundPage Not Found The content that you're looking for is unavailable. You might find what you are looking for by using the ...
Eswari Rao: Movies, Photos, Videos, News, Biography & Birthday | Times of IndiaWe use cookies and other tracking technologies to provide services while browsing the Website to show personalise conten...
What really went wrong for borders and angus & robertsonREDgroup Retail accounted for 20% of Australia’s $1.6B book market and in another life, might have been Australia’s vers...
404 - Page not foundHomeNG HindiIndiaIndiaAndhra PradeshArunachal PradeshAssamBiharChhattisgarhGoaGujaratHaryanaHimachal PradeshJharkhandKar...