Behavioral and neurophysiological aspects of working memory impairment in children with dyslexia
Behavioral and neurophysiological aspects of working memory impairment in children with dyslexia"
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT The present study aimed to identify behavioral and neurophysiological correlates of dyslexia which could potentially predict reading difficulty. One hundred and three Chinese
children with and without dyslexia (Grade 2 or 3, 7- to 11-year-old) completed both verbal and visual working memory (_n_-back) tasks with concurrent EEG recording. Data of 74 children with
sufficient usable EEG data are reported here. Overall, the typically developing control group (_N_ = 28) responded significantly faster and more accurately than the group with dyslexia (_N_
= 46), in both types of tasks. Group differences were also found in EEG band power in the retention phase of the tasks. Moreover, forward stepwise logistic regression demonstrated that both
behavioral and neurophysiological measures predicted reading difficulty uniquely. Dyslexia was associated with higher frontal midline theta activity and reduced upper-alpha power in the
posterior region. This finding is discussed in relation to the neural efficiency hypothesis. Whether these behavioral and neurophysiological patterns can longitudinally predict later reading
development among preliterate children requires further investigation. SIMILAR CONTENT BEING VIEWED BY OTHERS VOXEL AND SURFACE BASED WHOLE BRAIN ANALYSIS SHOWS READING SKILL ASSOCIATED
GREY MATTER ABNORMALITIES IN DYSLEXIA Article Open access 25 May 2021 BRAIN DYNAMICS OF (A)TYPICAL READING DEVELOPMENT—A REVIEW OF LONGITUDINAL STUDIES Article Open access 01 February 2021
DISRUPTED NETWORK INTERACTIONS SERVE AS A NEURAL MARKER OF DYSLEXIA Article Open access 03 November 2023 INTRODUCTION Children with developmental dyslexia have normal intelligence and
sufficient learning opportunities but experience unexpected difficulty in learning to read and write. Understanding the neurobiological basis of dyslexia might facilitate earlier and
possibly more accurate identification of children who will develop dyslexia, which is critical for early and most effective intervention. So far the most commonly observed brain differences
in people with dyslexia seem to be reduced activations in left temporal, parietal, and fusiform regions1. These studies mostly compared brain activations of people with and without dyslexia
during a variety of reading or reading-related tasks, such as fluent reading2 and phonological awareness tasks3. However, few studies have investigated brain activations of people with
dyslexia in a working memory (WM) task, although WM measures have been found to predict reading performance4. WM is a system that temporarily stores and manipulates information we need in
completing cognitive tasks5. In this model, visual and verbal information is maintained by domain-specific slave systems called the visuospatial sketch pad and the phonological loop, while
the central executive acts as an attentional-controlling component and coordinates the manipulation of information from the slave systems. Backward digit or word span tasks are widely used
to measure verbal WM, as this type of task requires participants to recall a sequence of digits or words in a reversed order from how they are presented (e.g., recalling “3–2–7” after
hearing “7–2–3”). Here participants need to not only maintain the original sequence of verbal information but also manipulate it to generate a new sequence in the reversed order.
Simultaneous storage and processing of information is a key characteristic of WM tasks, and essential for many cognitive skills. A review study has shown that there is a moderate correlation
between WM and academic achievement including both reading and math performance6. Below we briefly review previous studies linking reading development and WM. READING DEVELOPMENT AND WM WM
is closely related to the concept of short-term memory, which refers to the capacity to store information for a short time interval. Verbal short-term memory is typically measured by forward
span tasks that require participants to recall a sequence of digits or words in the same order as presented to them. Since there is no need to manipulate the information, short-term memory
tasks are considered to tap into part of WM functions only. In a meta-analytic review7, children with dyslexia performed significantly worse than chronological age controls on verbal
short-term memory tasks. However, when two other predictors (i.e., phonemic awareness and rime awareness) were controlled, verbal short-term memory did not contribute uniquely to children’s
word reading performance. Since WM measures reflect people’s ability to simultaneously maintain and process information, researchers have linked WM more frequently with reading
comprehension, a more complex reading ability that involves integrating information from earlier text, current input, and one’s long-term memory. A review study has reported a moderate
correlation between WM and reading comprehension4. Researchers have also investigated the association between WM and early reading development. For example, Chung and McBride-Chang8 found
that WM and inhibitory control together contributed uniquely to word reading performance of Hong Kong Chinese children at both the second and third years of kindergarten, after controlling
for age, vocabulary knowledge, and metalinguistic skills (see also the study of Welsh and colleagues9). These findings are consistent with a prevalent observation that individuals with
dyslexia tend to show deficits in WM10,11,12. Most reading studies have emphasized the role of verbal rather than visual working (or short-term) memory in predicting reading development.
This is because the majority of reading studies have been conducted among readers of alphabetic scripts, in which grapheme-phoneme correspondences allow the readers to decode the sound of a
spelling in a relatively easy manner13. However, the role of visual WM may be important in learning to read other types of scripts with a lack of grapheme-phoneme correspondences, e.g.,
Chinese. Since Chinese adopts a logographic writing system, children typically need to rely on rote character copying in Chinese literacy acquisition14. Hence, it is not surprising that
visual skills seem to be more important for Chinese readers than for alphabetic readers. In a cross-cultural study, Huang and Hanley15 found that performance on a task of visual paired
associates learning (i.e., learning the association between colors and abstract line drawings) was a strong predictor of the reading ability of 8-year-old primary children in Hong Kong and
Taiwan (Chinese script), but not in Britain (English script). Later studies also found that visual skills contributed uniquely to Chinese character reading after controlling for age,
vocabulary knowledge, and metalinguistic skills16. Although researchers have compared Chinese children with and without dyslexia on a variety of visual skill measures11 (e.g., visual search,
visual spatial relationship), few of them have adopted visual WM tasks. Therefore, the first aim of the present study was to fill in this research gap by using the _n_-back task17 with both
visual and verbal stimuli. In this task, participants are typically presented a sequence of items one by one and required to indicate whether the current item is the same as the one
presented _n_ items back. For example, the second “3” in the sequence “5–2–3–3–0” is a target in a 1-back task, and the second “2” in the sequence “9–2–0–2–3” is a target in a 2-back task.
As the sequence goes on, participants need to continuously update the information maintained in the WM and perform same-different judgment at the same time. WM demand varies with _n_ and
thus can be manipulated factorially. In the present study, two types of _n_-back task were used to compare Chinese children with and without dyslexia, one with Chinese characters as items
while the other with visual patterns. These two types of tasks would reflect children’s verbal and visual WM, respectively. Another important reason for choosing the _n_-back task is that
the procedure of this task fits the methodological constraints in many neuroimaging techniques, which is related to the second aim of the present study as spelled out below. BRAIN
OSCILLATIONS IN RELATION TO WM PROCESSES AND INDIVIDUAL DIFFERENCES Despite the close relation between WM and reading development, few studies have investigated the potential
neurophysiological substrate of a WM deficit in dyslexia. Nevertheless, the relation between brain oscillations and maintenance of information in WM has been extensively studied in the past
few decades. A recent review18 has shown that theta activity in the frontal midline region is most consistently associated with WM maintenance, especially in verbal WM tasks. The most
typical finding is an increase in frontal midline theta during a retention phase of the WM task relative to the baseline19. Among 15 EEG studies identified in the review that involved at
least two load levels of verbal WM, ten reported stepwise increase of theta with memory load20. On the other hand, theta increase has been less consistently observed in visual WM tasks
(around half of the reviewed studies). Due to its commonly observed increase at higher task difficulties, frontal midline theta is considered to reflect the mental effort engaged in the
task21. Hence, one may expect to see more pronounced theta increase in less apt individuals, since they probably need even more effort to complete a demanding task than more apt individuals
do. This prediction is consistent with the neural efficiency hypothesis22, which posits that better performing individuals show reduced (more efficient) neural activity in cognitive tasks.
In an EEG study by Maurer and colleagues23, healthy adults were required to memorize 2 or 4 unfamiliar symbols (low or high load) for 3.5 s and then to decide if a probe was one of the
presented symbols (i.e., a Sternberg task24). The participants’ behavioral performance decreased from the low load condition to the high load condition, while their frontal midline theta in
the retention phase increased. Moreover, the theta increase correlated significantly with the decrease in accuracy at the individual level, indicating that the more difficult a given task
seemed to an individual (as indicated by the larger decrease in accuracy), the larger increase in frontal midline theta was observed. Another study by Brzezicka and colleagues25 recorded
intracranial EEG while patients with epilepsy were performing a Sternberg task with 3 load levels (1, 2, or 3 pictures). Theta power in the retention phase increased with memory load in the
hippocampus but decreased in the dorsolateral prefrontal cortex (DLPFC). Furthermore, the faster one participant responded, the larger theta power decrease with memory load was found in the
DLPFC. Although the load effect on theta power was partly in different directions, both studies found that better performing participants showed a tendency of reduced theta power (smaller
increase or larger decrease with memory load) in the retention phase of a WM task. In addition to theta power, alpha power has also been found to change with WM load, although the direction
of alpha change was inconsistent. According to the review by Pavlov and Kotchoubey18, the ratio of studies on verbal WM that found alpha increase versus decrease was 4:1, and this ratio
became 3:2 for studies on visual WM. By dividing the alpha band into subbands (lower- and upper-alpha), some researchers found that upper-alpha activity tended to increase with memory load
while lower-alpha activity tended to decrease20,23. One explanation is that upper-alpha increase reflects inhibition of task-irrelevant regions in the retention phase and that lower-alpha
decrease reflects release of inhibition in task-relevant regions20,26. In terms of individual difference, Grabner and colleagues27 found that higher intelligence was associated with higher
upper-alpha power (i.e., lower brain activation) in WM tasks, consistent with the neural efficiency hypothesis22. THE PRESENT STUDY So far, much fewer studies have investigated WM-related
brain oscillations in children28, and almost none for children with dyslexia29. The present study recorded concurrent EEG while the children with and without dyslexia were doing the _n_-back
task, in order to compare not only behavioral performance but also brain oscillations of the two groups in a WM task. In addition to verbal WM, nonverbal stimuli were also included to tap
visual WM (considering the logographic nature of the Chinese writing system), which seemed to have received less attention in the reading literature. Based on the previous findings mentioned
above, we chose to examine theta power in the frontal midline region19,21 as well as lower- and upper-alpha power23 in the posterior region30,31. Moreover, logistic regression analysis was
conducted to find out whether these behavioral and neurophysiological measures were significant predictors of dyslexia32,33. We hypothesized that the children with dyslexia would show poorer
behavioral performance in the WM task. The neural efficiency hypothesis predicted that the children with dyslexia would manifest higher theta and lower alpha power (i.e., more brain
activation) during the retention phase of the task. RESULTS In the current design, Type of task (verbal, visual) and Load level (1-back, 2-back) were two within-participants factors, while
Group (control, dyslexic) was a between-participants factor. Since the control group had a significantly higher non-verbal intelligence than the group with dyslexia (_p_ = 0.014), ANCOVA was
conducted on each of the dependent variables in the _n_-back task with Intelligence as a covariate: (1) reaction time (RT, i.e., the time interval between stimulus offset and response) when
the target was correctly hit, (2) _d_ prime17, which was calculated based on the hit and false alarm rates, (3) log-transformed theta power in the frontal midline region, (4)
log-transformed lower-alpha power in the posterior region, and (5) log-transformed upper-alpha power in the posterior region. The restriction to the frontal midline region for theta analysis
was based on previous studies that showed a topographically more restricted effect for theta compared to alpha20,23, while alpha activity was relatively widespread and most prominent over
posterior areas30,34. _D_ PRIME Calculated based on the hit and false alarm rates, _d_ prime (_d'_) can be considered as an index reflecting how well one can differentiate targets from
non-targets (the higher _d'_, the better). Table 1 shows the mean _d'_ in each of the four conditions for each group. The ANCOVA showed that the main effect of Intelligence on
_d'_ was significant (_F_(1,71) = 6.30, _p_ = 0.014, _ƞ__p_2 = 0.08) while all interactions involving this covariate were non-significant (_p_s ≥ 0.150). The main effects of Type
(_F_(1,71) = 14.87, _p_ < 0.001, _ƞ__p_2 = 0.17), Load (_F_(1,71) = 5.23, _p_ = 0.025, _ƞ__p_2 = 0.07), and Group (_F_(1,71) = 15.80, _p_ < 0.001, _ƞ__p_2 = 0.18) were significant,
showing that the _d'_ was higher in the verbal tasks than in the visual tasks and also higher in the 1-back tasks than in the 2-back tasks. Besides, the control group demonstrated a
higher sensitivity to detect the targets in the WM tasks than the group with dyslexia. The Load × Group interaction (_F_(1,71) = 4.90, _p_ = 0.030, _ƞ__p_2 = 0.07) and the Type × Load ×
Group interaction (_F_(1,71) = 4.84, _p_ = 0.031, _ƞ__p_2 = 0.06) were significant, while all other interactions were non-significant (_p_s ≥ 0.369). The Load effect was comparable between
the two groups in the verbal tasks (− 0.54 vs. − 0.58, _p_ = 0.863, _d_ = 0.04). In the visual tasks, the control group showed a significantly larger Load effect than the group with dyslexia
(− 1.53 vs. − 0.77, _p_ = 0.001, _d_ = − 0.80), possibly because the group with dyslexia performed poorly in the visual 1-back task already. REACTION TIME The ANCOVA showed that the main
effect of Intelligence on RT and interactions involving this covariate were all non-significant (_p_s ≥ 0.434). The main effect of Group (_F_(1,71) = 12.82, _p_ = 0.001, _ƞ__p_2 = 0.15) and
the Type × Load × Group interaction (_F_(1,71) = 4.56, _p_ = 0.036, _ƞ__p_2 = 0.06) were significant, while all the other effects were non-significant (_p_s ≥ 0.395). The group with dyslexia
showed a slightly larger Load effect than the control group in the verbal tasks (241 vs. 162 ms, _p_ = 0.129, _d_ = 0.37). In the visual tasks, the Load effect of the group with dyslexia
was marginally smaller (77 vs. 215 ms, _p_ = 0.079, _d_ = − 0.43), possibly because this group responded quite slowly in the visual 1-back task already. Post-hoc _t_-tests showed shorter RT
in the control group than in the group with dyslexia for all the conditions (verbal 1-back: _p_ = 0.005, _d_ = − 0.69; verbal 2-back: _p_ < 0.001, _d_ = − 0.96; visual 1-back: _p_ <
0.001, _d_ = − 0.99) except the visual 2-back one (_p_ = 0.129, _d_ = − 0.34). FRONTAL MIDLINE THETA In the _n_-back task, the children needed to make same-different judgment for each
presented item and to maintain the newly updated items before seeing the next one. The mean RT in each condition ranged from 779 to 1173 ms. We can infer that the last 1-s interval of the
3.5-s fixation period fell into the retention phase of the _n_-back task, before which the encoding of new information and the same-different judgment were completed. Thus, band power during
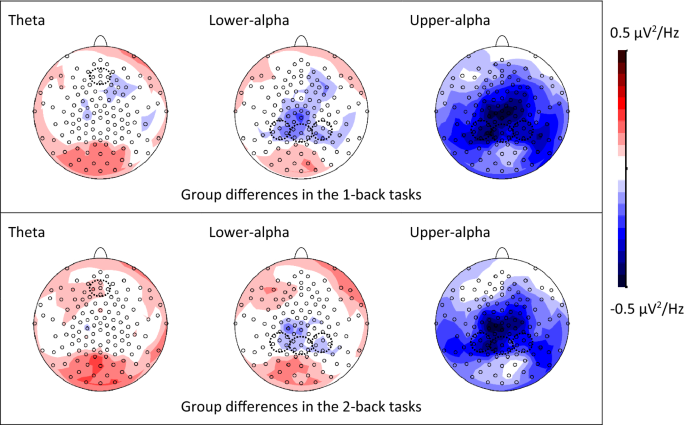
WM maintenance was obtained from this time window (see Fig. 1 and Fig. S1 for the topographic maps of band power differences between groups and between load levels, respectively). The
ANCOVA showed that the main effect of Intelligence on log-transformed theta power and interactions involving this covariate were all non-significant (_p_s ≥ 0.223). The Load × Group
interaction (_F_(1,71) = 5.99, _p_ = 0.017, _ƞ__p_2 = 0.08) was significant, while all the other effects were non-significant (_p_s ≥ 0.318). Post-hoc _t_-tests showed that the
log-transformed theta power of the control group had a marginally significant decrease from 1-back tasks to 2-back tasks (0.542 − 0.564 = − 0.022, _p_ = 0.078, _d_ = − 0.35). In contrast,
the group with dyslexia demonstrated an opposite trend (0.588 − 0.575 = 0.013, _p_ = 0.173, _d_ = 0.20). In terms of group difference, the group with dyslexia had a non-significantly higher
theta power than the control group in both 1-back and 2-back tasks (1-back: 0.575 − 0.564 = 0.011, _p_ = 0.818, _d_ = 0.06; 2-back: 0.588 − 0.542 = 0.046, _p_ = 0.325, _d_ = 0.24).
UPPER-ALPHA IN THE POSTERIOR REGION The ANCOVA showed that the main effect of Intelligence on log-transformed upper-alpha power and interactions involving this covariate were all
non-significant (_p_s ≥ 0.100). The main effect of Group (_F_(1,71) = 4.91, _p_ = 0.030, _ƞ__p_2 = 0.07) was significant, while all the other effects were non-significant (_p_s ≥ 0.188). The
control group demonstrated a higher upper-alpha power in the posterior region than the group with dyslexia. LOWER-ALPHA IN THE POSTERIOR REGION The ANCOVA showed that all the effects were
non-significant on log-transformed lower-alpha power (_p_s ≥ 0.119). DISTINGUISHING CHILDREN WITH AND WITHOUT DYSLEXIA To find out which variables were most strongly and uniquely associated
with dyslexia, the following variables from the WM tasks were entered by block32 into logistic regression models as predictors of dyslexia: (1) Block 1 included general control variables:
age, grade, non-verbal intelligence; (2) Block 2 included 8 behavioral variables: reaction time and _d_ prime in each of the four conditions; and (3) Block 3 included 12 neurophysiological
variables: log-transformed frontal midline theta, posterior lower- and upper-alpha power in each of the four conditions. The forward Wald method33 was adopted in each block so as to identify
behavioral measures (if any) that uniquely predicted dyslexia beyond control variables as well as unique neurophysiological predictors (if any) beyond both control and behavioral variables.
Table 2 shows the three logistic regression models generated in each block. In Block 1, the Wald statistic showed that only non-verbal intelligence was a significant predictor (_p_ =
0.040). Model 1 made significantly better prediction of dyslexia than a null model (_χ_2(1) = 4.99, _p_ = 0.025). In Block 2, two behavioral variables (i.e., RT in the visual 1-back
condition, _d'_ in the verbal 2-back condition) were entered (_p_s ≤ 0.009), and Intelligence became non-significant (_p_ = 0.328). Model 2 significantly improved prediction of dyslexia
relative to Model 1 (_χ_2(2) = 24.53, _p_ < 0.001). The classification accuracy increased from 59.5% (dyslexic: 82.6%; control: 21.4%) to 81.1% (dyslexic: 84.8%; control: 75.0%), and
Nagelkerke’s _R_2 improved from 0.089 to 0.448. In Block 3, two neurophysiological variables (i.e., log-transformed frontal midline theta in the verbal 2-back condition, log-transformed
posterior upper-alpha in the visual 1-back condition) were further entered (_p_s ≤ 0.006), while the two behavioral variables remained significant (_p_s ≤ 0.002). Model 3 significantly
improved prediction of dyslexia relative to Model 2 (_χ_2(2) = 16.27, _p_ < 0.001). The classification accuracy further increased from 81.1 to 82.4% (dyslexic: 87.0%; control: 75.0%).
Nagelkerke’s _R_2 of Model 3 was 0.628, suggesting a relatively strong relation between the predictors and dyslexia (see Fig. S2 for the distribution of predicted probabilities in the
classification plots). To sum up, significant and unique predictors of dyslexia were found from both behavioral and neurophysiological measures of WM in the present study. Figure 2 displays
the scatterplots of the two groups, showing each significant predictor of dyslexia (y axis) as a function of non‐verbal intelligence (x axis). To further examine the predictive role of WM
measures in dyslexia without any confounding effect of non‐verbal intelligence, we conducted additional logistic regression analysis on the control group and a subsample of the group with
dyslexia (i.e., excluding those who scored 21 or lower in non‐verbal intelligence), whose non‐verbal intelligence scores were comparable (_p_ = 0.860). The results showed that non‐verbal
intelligence was no longer a significant predictor while behavioral and neurophysiological measures of WM (i.e., verbal 2-back RT, visual 1-back _d'_, verbal 2-back theta and
upper-alpha) still significantly and uniquely predicted dyslexia. More details of the additional logistic regression analysis can be found in the Supplemental Material. DISCUSSION The
present study required Chinese children with and without dyslexia to complete verbal and visual WM tasks with concurrent EEG recording. Consistent with the findings of previous
studies10,11,12, the control group responded significantly faster and more accurately than the group with dyslexia, in both types of tasks. In the retention phase of the tasks, the frontal
midline theta power of the control group tended to decrease from the low load to the high load condition, while the group with dyslexia showed an opposite trend. The control group also
demonstrated a higher upper-alpha power than the group with dyslexia did across all conditions. Forward stepwise logistic regression identified a few significant predictors of dyslexia,
including both behavioral and neurophysiological measures of WM. WM-RELATED NEUROPHYSIOLOGICAL CORRELATES OF DYSLEXIA In the logistic regression model, frontal midline theta activity in the
verbal 2-back task was positively associated with dyslexia, and posterior upper-alpha power in the visual 1-back task was negatively associated with dyslexia. This means that children in our
sample who spent more mental effort and inhibited brain activation to a lesser extent during WM maintenance tended to have dyslexia, which is consistent with the neural efficiency
hypothesis22. Importantly, these neurophysiological correlates uniquely predicted reading difficulty in addition to non-verbal intelligence (though non-significant in the final model) and
behavioral correlates. While behavioral indicators (speed and accuracy) reflected the children’s final achievement in WM tasks, neurophysiological measures (theta and alpha oscillations)
reflected the amount of mental effort and neural resources engaged during WM maintenance21,23. These variables tapped into different aspects of WM, and all predicted reading difficulty
uniquely. A few fMRI studies have also compared brain activation of dyslexic and non-dyslexic readers during WM tasks. While some found reduced prefrontal activation in dyslexia35, some
found both increased and decreased activation in the prefrontal cortex36. These mixed results could be partly due to the differences in the tasks. Researchers have found that neural
efficiency could be modulated by task difficulty and other factors37. There are situations where better performing individuals may display the same amount of or even more brain activation38.
Besides, the relatively low temporal resolution of the fMRI technique could be another reason. An EEG study by Jaušovec and Jaušovec39 showed that in a modified _n_-back task, the theta
power of high-intelligence individuals was higher than that of low-intelligence ones in 0–500 ms post-stimulus onset. Then it decreased soon and the group difference was reversed in
1000–2000 ms, suggesting that the high-intelligence individuals were more intensely engaged with the task in the first 500 ms and soon lowered their mental effort in the following retention
phase. Hence, the retention phase, the phase selected for analysis in the present study, might be an optimal time interval for reduced neural activation in better performing individuals
(i.e., neural efficiency) to occur. Previous fMRI studies might have captured neural activation across different phases, which could be one of the reasons for the mixed findings. By using
EEG, the present study was able to examine brain activation in the retention phase more precisely. In contrast to the prominent Group effect, the Load effect on brain oscillations seemed to
be relatively minor in the present study. For frontal midline theta power, only the Load × Group interaction was significant, caused by the opposite directions of theta change in the two
groups with increasing load. While the control group had a marginally significant theta decrease, the group with dyslexia demonstrated a non-significant trend of theta increase. For lower-
and upper-alpha power, no significant difference was found between load levels. Different patterns of theta and alpha changes with WM load have been reported in previous studies, and the
modulating factors remain largely unclear18. For example, a few studies found an inverted U-shape influence of WM load on frontal midline theta power, i.e., highest theta power with a
moderate load level40,41. So, one explanation for the trend of theta decrease in the control group could be that the current 2-back tasks were beyond the moderate load level for early grade
children. However, this account would predict a theta decrease in the group with dyslexia as well, since their behavioral performance was even worse than that of the control group. This
prediction contradicted the observed trend of theta increase in the group with dyslexia. Note that the Load effect was not significant in either group. Further studies are needed to examine
the robustness of the observed theta change when children perform WM tasks with different load levels. WM IMPAIRMENT: CAUSE OR EFFECT The present study identified a few behavioral and
neurophysiological correlates (i.e., visual 1-back RT and upper-alpha, verbal 2-back _d'_ and theta) of WM impairment in dyslexia (i.e., more neural resources engaged but poorer
behavioral performance). However, we cannot tell whether poor WM is a cause or an effect of dyslexia. In the verbal _n_-back tasks, the children were able to make use of phonological codes
of the Chinese characters to achieve better performance than in the visual tasks, where only visual codes were usable. The group differences in the verbal tasks could result from inefficient
processing of Chinese characters in the children with dyslexia, due to their poor orthography–phonology conversion42. Hence, poor verbal WM could be a consequence of other deficits in
dyslexia. On the other hand, similar group differences were observed in the visual _n_-back tasks, where non-verbal stimuli were used without involving orthographic or phonological encoding.
The visual 1-back RT and upper-alpha were also unique predictors of dyslexia in the logistic regression model. This finding seems to support a deficit in the central executive component of
WM43,44, whose dysfunction impairs both verbal and visual working memories. Nevertheless, an alternative explanation is that the development of visual processing is influenced by the
children’s reading skills45. In a longitudinal study, Pan and colleagues42 tracked Chinese children’s character reading accuracy and pure visual skill for three years (at ages 6 to 8). They
found that reading accuracy predicted subsequent performance in a pure visual task but not vice versa. Hence, poor visual WM could be a consequence of reading difficulty as well. Despite
these alternative possibilities, we believe that the children with dyslexia are highly likely to have central executive dysfunction given the available evidence. Melby-Lervåg and colleagues7
have shown that verbal short-term memory does not contribute uniquely to word reading performance when metalinguistic skills are controlled. But WM together with other executive functions
seems to have a unique contribution to reading achievement8,9,46, suggesting the importance of central executive in reading development. In the present study, similar group differences were
observed across the two types of WM tasks, consistent with the existence of central executive dysfunction in dyslexia. Note that developmental dyslexia can be classified into different
subtypes47,48, so not all children with dyslexia have the same deficits or the same cause of the deficits. The scatterplots in Fig. 2 show that not all children in the dyslexic group
appeared to have WM impairment and that some control children demonstrated poor WM. Although WM differences can be observed in between-groups comparisons, WM impairment does not always
co-occur with dyslexia49. In the current logistic regression analysis, Model 3 only correctly classified 87.0% of the children with dyslexia and 75.0% of the control children. Hence,
behavioral and neurophysiological measures of WM could potentially predict dyslexia, but these measures alone are insufficient to classify children as having dyslexia or not. The present
study adopted the _n_-back task and identified both behavioral and neurophysiological correlates of WM impairment in dyslexia, among early grade children. Importantly, EEG band power (theta
and alpha oscillations) uniquely predicted reading difficulty in addition to the behavioral measures (RT and _d'_). One limitation of the present study is that the non-verbal
intelligence of the two groups was not matched, although additional logistic regression analysis on the subsample showed a similar pattern of results after matching the non‐verbal
intelligence scores. Besides, previous studies found that dyslexic children’s deficits in certain short-term memory tasks disappeared when they were matched to controls on non-verbal
intelligence and oral language abilities50,51. Although the current _n_-back tasks required key-pressing responses only and involved the use of oral language minimally (especially the visual
tasks), this study could be improved by including measures of oral language abilities. With stricter control on non-verbal intelligence and oral language abilities, future studies may
replicate the present study among younger or preliterate children and follow them up52 to find out whether earlier behavioral and neurophysiological measures of WM longitudinally predict
later reading development. Compared to the backward span task and some other WM tasks, the _n_-back task has at least two advantages. First, the children are able to respond by simply
pressing a key without verbal articulation, so that any confounding effects of language abilities are minimized. Second, the procedure of this task allows concurrent EEG recording and
frequency analysis in a retention phase, which could potentially improve the prediction success. Longitudinal studies are needed to examine the effectiveness of these potential predictors
and to determine whether they can be used in clinical practice. CONCLUSIONS In the 1-back and 2-back WM tasks, the typically developing children performed better than the children with
dyslexia. Frontal midline theta and posterior upper-alpha power in the retention phase of the tasks reflected the amount of mental effort and neural resources being engaged, and they
predicted dyslexia uniquely in addition to indices of speed and accuracy. However, it remains unclear whether these behavioral and neurophysiological patterns are merely consequences of
reading difficulty or not. Further investigation is needed to examine whether the current measures can be used to predict reading difficulty in pre-readers. METHODS PARTICIPANTS Data of 28
typically reading children and 46 children with dyslexia (native Cantonese-speaking; second or third grade; 7–11 years old) from a larger research project (approved by The Joint Chinese
University of Hong Kong—New Territories East Cluster Clinical Research Ethics Committee) are reported. They were recruited from Hong Kong primary schools and education authorities, and
written informed consent was obtained from the children and their guardians. All methods were performed in accordance with relevant guidelines. Among 103 children (66 with dyslexia) who
completed all the relevant tasks as described below, 29 children (20 with dyslexia) were excluded from the present study due to too few usable EEG segments (see section on EEG recording and
preprocessing below). The typically developing children had no difficulty in reading or writing based on parents’ report. Those with dyslexia were formally diagnosed by either educational or
clinical psychologists based on The Hong Kong Test of Specific Learning Difficulties in Reading and Writing for Primary School Students—Third Edition [HKT-P(III)]53, whose criteria included
adequate IQ (higher than 85), poor literacy (− 1 _SD_ or below), and at least one area of cognitive-linguistic deficit (− 1 _SD_ or below)54. Besides, they had no history of significant
sensory impairment, birth complications, or brain injury. Based on the parents’ responses in a questionnaire (_N_ = 70), 7 children from the dyslexic group (16.7%) and 2 from the control
group (7.1%) had a formal diagnosis of language impairment (_p_ = 0.244). Table 3 shows the demographic information of the two groups, who did not differ significantly in gender, age, grade,
maternal or paternal education level, or monthly family income (_p_s ≥ 0.248). PROCEDURE RAVEN’S STANDARD PROGRESSIVE MATRIX (RSPM) RSPM55, Sets A to C were used to assess non-verbal
intelligence. Each test item required the children to choose an option out of six (Sets A and B) or eight (Set C) to fill the missing part of a design. There were 12 test items in each set
and thus 36 items in total. VERBAL N-BACK TASK Sixty Grade-2 level characters with a varying number of strokes (from 4 to 13) were selected from the Hong Kong Corpus of Primary School
Chinese56 and formed a sequence for verbal 1-back and 2-back tasks respectively. In each task, twenty of the characters appeared twice, and those appearing at the second time were targets.
The targets appeared immediately after the same characters in the 1-back task and appeared the second after the same characters in the 2-back task (Fig. 3). The same set of characters were
used in both tasks, but the target characters were mostly different so that one could not predict whether a character was a target or not based on its status in an earlier task. Each task
contained 60 non-targets and 20 targets and was divided into two blocks of 40 characters. The characters were sequenced in a way that there was no obvious semantic relatedness or
orthographic similarity between each pair of consecutive characters, except for the targets in the 1-back task. The _n_-back task was administered with the E-Prime 3.0 software
(https://pstnet.com/products/e-prime/). Each character appeared for 500 ms, followed by a 3500-ms fixation and then the next character. The children needed to press “1” on the keyboard as
accurately and quickly as possible, when they detected a target, and did not need to press any key for non-targets. At the beginning of each task, the task requirement (1-back or 2-back) was
explained to the children, and they needed to complete a practice block with 14 characters (4 targets) to make sure that they understood the task requirement. They were able to repeat the
practice once or more if needed. VISUAL N-BACK TASK Thirty 3 × 3 checkerboard patterns were created for visual 1-back and 2-back tasks. Each pattern contained 3 black squares and 6 white
ones (see Supplemental Material). We did not use 60 different patterns, as it would yield too many similar patterns making the task too difficult. Similar to the verbal _n_-back task, both
visual 1-back and 2-back tasks consisted of two blocks of 40 items (30 non-targets and 10 targets each). The patterns were sequenced in a way that each pair of consecutive patterns did not
look similar, except for the targets in the 1-back task. The procedure was the same as that of the verbal _n_-back task. Each child completed all the four tasks (2 types × 2 load levels),
whose order was counterbalanced across participants for each group. EEG RECORDING AND PREPROCESSING EEG data were recorded at 500 Hz (online filter: 0.1–100 Hz; Cz as recording reference)
using an EGI (Electrical Geodesics, Inc.) 128-channel system while each child was completing the _n_-back task in a quiet room. Impedances were controlled below 50 kΩ. The EEG recordings
were offline filtered (0.1–70 Hz; notch filter: 50 Hz). Bad channels were excluded, and the remaining data were submitted to independent component analysis for eye movement correction57. The
excluded channels were then spline interpolated58. All data were re-referenced to the average reference59 and then segmented to include the last 1-s interval of the fixation period (i.e.,
before the onset of the next item; see Fig. 4). Only segments following non-targets (without motor artefacts from key pressing) and not exceeding ± 100 μV were used in further analysis.
Twenty-nine children with poor EEG data quality in any of the four conditions were excluded from the present study, so that all the 74 children reported here had no fewer than 10 usable
epochs in each condition. The average number of usable EEG epochs was 29, 28, 31, and 29 in the verbal 1-back, verbal 2-back, visual 1-back, and visual 2-back conditions respectively. Before
the _n_-back task, EEG recordings were also obtained during a 3-min eyes-closed resting state block, with the same setting. The resting EEG data went through similar preprocessing steps and
were segmented into 1-s epochs for determination of individual alpha frequency60 (IAF). The fast Fourier transformation was applied to each of the EEG epochs obtained from the _n_-back task
and the eyes-closed state. For each child, the frequency with peak power density in the alpha band (8–13 Hz) across all electrodes in the eyes-closed state was identified as IAF. For two
children, the alpha peak failed to be detected with power density averaged across electrodes, and power density at the occipital electrode Oz was then used to identify IAF; for another two
children with eyes-closed EEG data of poor quality, their IAF was set at 10. The two groups did not differ significantly in IAF (_p_ = 0.438). Three frequency bands were defined with IAF as
an individual anchor point: (IAF-6) to (IAF-3) as theta band, (IAF-2) to (IAF-1) as lower-alpha band, and IAF to (IAF + 1) as upper-alpha band60. For each child, mean theta power in the
frontal midline region (pooled from AFz plus 4 surrounding electrodes) was calculated for each of the four conditions separately and log-transformed to approach a normal distribution across
children23,61. Mean power in the lower- and upper-alpha bands in the posterior region30,31 (pooled from Pz, P3, P4 and 15 surrounding electrodes) was calculated and log-transformed as well
for each condition and child. DATA AVAILABILITY The data that support the findings of this study are available from the corresponding author upon reasonable request. REFERENCES * Norton, E.
S., Beach, S. D. & Gabrieli, J. D. E. Neurobiology of dyslexia. _Curr. Opin. Neurobiol._ 30, 73–78 (2015). Article CAS PubMed Google Scholar * Langer, N., Benjamin, C., Minas, J.
& Gaab, N. The neural correlates of reading fluency deficits in children. _Cereb. Cortex_ 25, 1441–1453 (2015). Article PubMed Google Scholar * Kovelman, I. _et al._ Brain basis of
phonological awareness for spoken language in children and its disruption in dyslexia. _Cereb. Cortex_ 22, 754–764 (2012). Article PubMed Google Scholar * Follmer, D. J. Executive
function and reading comprehension: A meta-analytic review. _Educ. Psychol._ 53, 42–60 (2018). Article Google Scholar * Baddeley, A. Working memory. _Science_ 255, 556–559 (1992). Article
ADS CAS PubMed Google Scholar * Jacob, R. & Parkinson, J. The potential for school-based interventions that target executive function to improve academic achievement: A review.
_Rev. Educ. Res._ 85, 512–552 (2015). Article Google Scholar * Melby-Lervåg, M., Lyster, S.-A.H. & Hulme, C. Phonological skills and their role in learning to read: A meta-analytic
review. _Psychol. Bull._ 138, 322–352 (2012). Article PubMed Google Scholar * Chung, K. K. H. & McBride-Chang, C. Executive functioning skills uniquely predict Chinese reading. _J.
Educ. Psychol._ 103, 909–921 (2011). Article Google Scholar * Welsh, J. A., Nix, R. L., Blair, C., Bierman, K. L. & Nelson, K. E. The development of cognitive skills and gains in
academic school readiness for children from low-income families. _J. Educ. Psychol._ 102, 43–53 (2010). Article PubMed PubMed Central Google Scholar * Chiappe, P., Siegel, L. S. &
Hasher, L. Working memory, inhibitory control, and reading disability. _Mem. Cogn._ 28, 8–17 (2000). Article CAS Google Scholar * Peng, P., Wang, C., Tao, S. & Sun, C. The deficit
profiles of Chinese children with reading difficulties: A meta-analysis. _Educ. Psychol. Rev._ 29, 513–564 (2017). Article Google Scholar * Reiter, A., Tucha, O. & Lange, K. W.
Executive functions in children with dyslexia. _Dyslexia_ 11, 116–131 (2005). Article PubMed Google Scholar * Gibson, E. J., Pick, A., Osser, H. & Hammond, M. The role of
grapheme-phoneme correspondence in the perception of words. _Am. J. Psychol._ 75, 554–570 (1962). Article CAS PubMed Google Scholar * McBride-Chang, C., Chung, K. K. H. & Tong, X.
Copying skills in relation to word reading and writing in Chinese children with and without dyslexia. _J. Exp. Child Psychol._ 110, 422–433 (2011). Article PubMed Google Scholar * Huang,
H. S. & Hanley, J. R. Phonological awareness and visual skills in learning to read Chinese and English. _Cognition_ 54, 73–98 (1995). Article CAS PubMed Google Scholar * Liu, D.,
Chen, X. & Chung, K. K. H. Performance in a visual search task uniquely predicts reading abilities in third-grade Hong Kong Chinese children. _Sci. Stud. Read._ 19, 307–324 (2015).
Article Google Scholar * Haatveit, B. C. _et al._ The validity of d prime as a working memory index: Results from the ‘Bergen n-back task’. _J. Clin. Exp. Neuropsychol._ 32, 871–880
(2010). Article PubMed Google Scholar * Pavlov, Y. G. & Kotchoubey, B. Oscillatory brain activity and maintenance of verbal and visual working memory: A systematic review.
_Psychophysiology_ 59, e13735 (2020). PubMed Google Scholar * Onton, J., Delorme, A. & Makeig, S. Frontal midline EEG dynamics during working memory. _Neuroimage_ 27, 341–356 (2005).
Article PubMed Google Scholar * Michels, L., Moazami-Goudarzi, M., Jeanmonod, D. & Sarnthein, J. EEG alpha distinguishes between cuneal and precuneal activation in working memory.
_Neuroimage_ 40, 1296–1310 (2008). Article PubMed Google Scholar * Gevins, A., Smith, M. E., McEvoy, L. & Yu, D. High-resolution EEG mapping of cortical activation related to working
memory: Effects of task difficulty, type of processing, and practice. _Cereb. Cortex_ 7, 374–385 (1997). Article CAS PubMed Google Scholar * Haier, R. J. _et al._ Cortical glucose
metabolic rate correlates of abstract reasoning and attention studied with positron emission tomography. _Intelligence_ 12, 199–217 (1988). Article Google Scholar * Maurer, U. _et al._
Frontal midline theta reflects individual task performance in a working memory task. _Brain Topogr._ 28, 127–134 (2015). Article PubMed Google Scholar * Sternberg, S. High-speed scanning
in human memory. _Science_ 153, 652–654 (1966). Article ADS CAS PubMed Google Scholar * Brzezicka, A. _et al._ Working memory load-related theta power decreases in dorsolateral
prefrontal cortex predict individual differences in performance. _J. Cogn. Neurosci._ 31, 1290–1307 (2019). Article PubMed PubMed Central Google Scholar * Klimesch, W., Sauseng, P. &
Hanslmayr, S. EEG alpha oscillations: The inhibition–timing hypothesis. _Brain Res. Rev._ 53, 63–88 (2007). Article PubMed Google Scholar * Grabner, R. H., Fink, A., Stipacek, A.,
Neuper, C. & Neubauer, A. C. Intelligence and working memory systems: Evidence of neural efficiency in alpha band ERD. _Cogn. Brain Res._ 20, 212–225 (2004). Article CAS Google Scholar
* Güntekin, B. _et al._ Theta and alpha oscillatory responses differentiate between six-to seven-year-old children and adults during successful visual and auditory memory encoding. _Brain
Res._ 1747, 147042 (2020). Article PubMed CAS Google Scholar * Martínez-Briones, B. J., Fernández-Harmony, T., Garófalo Gómez, N., Biscay-Lirio, R. J. & Bosch-Bayard, J. Working
memory in children with learning disorders: An EEG power spectrum analysis. _Brain Sci._ 10, 817 (2020). Article PubMed Central Google Scholar * Hu, Z. _et al._ Working memory capacity is
negatively associated with memory load modulation of alpha oscillations in retention of verbal working memory. _J. Cogn. Neurosci._ 31, 1933–1945 (2019). Article PubMed PubMed Central
Google Scholar * Sander, M. C., Werkle-Bergner, M. & Lindenberger, U. Amplitude modulations and inter-trial phase stability of alpha-oscillations differentially reflect working memory
constraints across the lifespan. _Neuroimage_ 59, 646–654 (2012). Article PubMed Google Scholar * Kraft, I. _et al._ Predicting early signs of dyslexia at a preliterate age by combining
behavioral assessment with structural MRI. _Neuroimage_ 143, 378–386 (2016). Article PubMed Google Scholar * Puolakanaho, A. _et al._ Very early phonological and language skills:
Estimating individual risk of reading disability. _J. Child Psychol. Psychiatry_ 48, 923–931 (2007). Article PubMed Google Scholar * Jensen, O., Gelfand, J., Kounios, J. & Lisman, J.
E. Oscillations in the alpha band (9–12 Hz) increase with memory load during retention in a short-term memory task. _Cereb. Cortex_ 12, 877–882 (2002). Article PubMed Google Scholar *
Beneventi, H., Tønnessen, F. E., Ersland, L. & Hugdahl, K. Executive working memory processes in dyslexia: Behavioral and fMRI evidence. _Scand. J. Psychol._ 51, 192–202 (2010). Article
PubMed Google Scholar * Vasic, N., Lohr, C., Steinbrink, C., Martin, C. & Wolf, R. C. Neural correlates of working memory performance in adolescents and young adults with dyslexia.
_Neuropsychologia_ 46, 640–648 (2008). Article PubMed Google Scholar * Neubauer, A. C. & Fink, A. Intelligence and neural efficiency. _Neurosci. Biobehav. Rev._ 33, 1004–1023 (2009).
Article PubMed Google Scholar * Larson, G. E., Haier, R. J., LaCasse, L. & Hazen, K. Evaluation of a “mental effort” hypothesis for correlations between cortical metabolism and
intelligence. _Intelligence_ 21, 267–278 (1995). Article Google Scholar * Jaušovec, N. & Jaušovec, K. Differences in induced brain activity during the performance of learning and
working-memory tasks related to intelligence. _Brain Cogn._ 54, 65–74 (2004). Article PubMed Google Scholar * Gärtner, M., Rohde-Liebenau, L., Grimm, S. & Bajbouj, M. Working
memory-related frontal theta activity is decreased under acute stress. _Psychoneuroendocrinology_ 43, 105–113 (2014). Article PubMed Google Scholar * Zhang, D., Zhao, H., Bai, W. &
Tian, X. Functional connectivity among multi-channel EEGs when working memory load reaches the capacity. _Brain Res._ 1631, 101–112 (2016). Article CAS PubMed Google Scholar * Pan, J.,
Cui, X., McBride, C. & Shu, H. An investigation of the bidirectional relations of word reading to timed visual tasks involving different levels of phonological processing in Chinese.
_Sci. Stud. Read._ 24, 275–291 (2020). Article Google Scholar * Smith-Spark, J. H. & Fisk, J. E. Working memory functioning in developmental dyslexia. _Memory_ 15, 34–56 (2007).
Article PubMed Google Scholar * Swanson, H. L. & Sachse-Lee, C. A subgroup analysis of working memory in children with reading disabilities: Domain-general or domain-specific
deficiency?. _J. Learn. Disabil._ 34, 249–263 (2001). Article CAS PubMed Google Scholar * McBride-Chang, C. _et al._ Visual spatial skill: A consequence of learning to read?. _J. Exp.
Child Psychol._ 109, 256–262 (2011). Article PubMed Google Scholar * Liu, C., Chung, K. K. H. & Fung, W. K. Bidirectional relationships between children’s executive functioning,
visual skills, and word reading ability during the transition from kindergarten to primary school. _Contemp. Educ. Psychol._ 59, 101779 (2019). Article Google Scholar * Ho, C.S.-H., Chan,
D. W., Chung, K. K. H., Lee, S.-H. & Tsang, S.-M. In search of subtypes of Chinese developmental dyslexia. _J. Exp. Child Psychol._ 97, 61–83 (2007). Article PubMed Google Scholar *
Wang, L.-C. & Yang, H.-M. Classifying Chinese children with dyslexia by dual-route and triangle models of Chinese reading. _Res. Dev. Disabil._ 35, 2702–2713 (2014). Article PubMed
Google Scholar * Gray, S. _et al._ Working memory profiles of children with dyslexia, developmental language disorder, or both. _J. Speech Lang. Hear. Res._ 62, 1839–1858 (2019). Article
PubMed PubMed Central Google Scholar * Cowan, N. _et al._ Short-term memory in childhood dyslexia: Deficient serial order in multiple modalities. _Dyslexia_ 23, 209–233 (2017). Article
PubMed PubMed Central Google Scholar * Rispens, J. & Baker, A. Nonword repetition: The relative contributions of phonological short-term memory and phonological representations in
children with language and reading impairment. _J. Speech Lang. Hear. Res._ 55, 683–694 (2012). Article PubMed Google Scholar * Carroll, J. M., Solity, J. & Shapiro, L. R. Predicting
dyslexia using prereading skills: The role of sensorimotor and cognitive abilities. _J. Child Psychol. Psychiatry_ 57, 750–758 (2016). Article PubMed Google Scholar * Ho, C.S.-H. _et al._
_The Hong Kong Test of Specific Learning Difficulties in Reading and Writing for Primary School Students_ 3rd edn. (Hong Kong Specific Learning Difficulties Research Team, 2016). Google
Scholar * Chung, K. K. H. Understanding developmental dyslexia in Chinese: Linking research to practice. _Asia Pac. J. Dev. Differ._ 4, 3–15 (2017). Article Google Scholar * Raven, J. The
Raven’s Progressive Matrices: Change and stability over culture and time. _Cogn. Psychol._ 41, 1–48 (2000). Article CAS PubMed Google Scholar * Leung, M. T. & Lee, A. _The Hong Kong
corpus of primary school Chinese_. Paper presented at the ninth meeting of the International Clinical Phonetics and Linguistics Association, Hong Kong, China (2002). * Jung, T.-P. _et al._
Removal of eye activity artifacts from visual event-related potentials in normal and clinical subjects. _Clin. Neurophysiol._ 111, 1745–1758 (2000). Article CAS PubMed Google Scholar *
Perrin, F., Pernier, J., Bertnard, O., Giard, M. H. & Echallier, J. F. Mapping of scalp potentials by surface spline interpolation. _Electroencephalogr. Clin. Neurophysiol._ 66, 75–81
(1987). Article CAS PubMed Google Scholar * Lehmann, D. & Skrandies, W. Reference-free identification of components of checkerboard-evoked multichannel potential fields.
_Electroencephalogr. Clin. Neurophysiol._ 48, 609–621 (1980). Article CAS PubMed Google Scholar * Doppelmayr, M., Klimesch, W., Pachinger, T. & Ripper, B. Individual differences in
brain dynamics: Important implications for the calculation of event-related band power. _Biol. Cybern. Heidelb._ 79, 49–57 (1998). Article CAS Google Scholar * Gasser, T., Bächer, P.
& Möcks, J. Transformations towards the normal distribution of broad band spectral parameters of the EEG. _Electroencephalogr. Clin. Neurophysiol._ 53, 119–124 (1982). Article CAS
PubMed Google Scholar Download references ACKNOWLEDGEMENTS This research was supported by the Health and Medical Research Fund (HMRF) of the Food and Health Bureau, Hong Kong (04152496).
AUTHOR INFORMATION Author notes * These authors contributed equally: Jie Wang and Shuting Huo. AUTHORS AND AFFILIATIONS * Department of Psychology, The Education University of Hong Kong,
Hong Kong S.A.R., China Jie Wang * Department of Psychology, The Chinese University of Hong Kong, Hong Kong S.A.R., China Shuting Huo, Ka Chun Wu, Jianhong Mo, Wai Leung Wong & Urs
Maurer * Brain and Mind Institute, The Chinese University of Hong Kong, Hong Kong S.A.R., China Shuting Huo & Urs Maurer * Department of Educational Psychology, The Chinese University of
Hong Kong, Hong Kong S.A.R., China Ka Chun Wu Authors * Jie Wang View author publications You can also search for this author inPubMed Google Scholar * Shuting Huo View author publications
You can also search for this author inPubMed Google Scholar * Ka Chun Wu View author publications You can also search for this author inPubMed Google Scholar * Jianhong Mo View author
publications You can also search for this author inPubMed Google Scholar * Wai Leung Wong View author publications You can also search for this author inPubMed Google Scholar * Urs Maurer
View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS U.M. designed the study. K.C.W., J.M., W.L.W., and J.W. prepared and conducted the study.
S.H. and J.W. analyzed the data. J.W. wrote the paper. U.M., S.H., and J.W. edited the manuscript. All authors reviewed the manuscript. CORRESPONDING AUTHOR Correspondence to Urs Maurer.
ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. ADDITIONAL INFORMATION PUBLISHER'S NOTE Springer Nature remains neutral with regard to jurisdictional
claims in published maps and institutional affiliations. SUPPLEMENTARY INFORMATION SUPPLEMENTARY INFORMATION. RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative
Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the
original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in
the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your
intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence,
visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Wang, J., Huo, S., Wu, K.C. _et al._ Behavioral and neurophysiological
aspects of working memory impairment in children with dyslexia. _Sci Rep_ 12, 12571 (2022). https://doi.org/10.1038/s41598-022-16729-8 Download citation * Received: 21 January 2022 *
Accepted: 14 July 2022 * Published: 22 July 2022 * DOI: https://doi.org/10.1038/s41598-022-16729-8 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this
content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative
Trending News
In vivo CRISPR inactivation of Fos promotes prostate cancer progression by altering the associated AP-1 subunit JunProstate cancer is a major global health concern with limited treatment options for advanced disease. Its heterogeneity ...
9 best gardening gifts for the green-fingered person in your lifeThere are heaps of potential gifts out there for the gardener in your life, but not all will be greeted with a bright su...
Marilyn manson dropped by record label after evan rachel wood's abuse allegationsMarilyn Manson has been dropped by his record label after Evan Rachel Wood and other women accused the musician and acto...
Just a moment...Central Dentists Tingalpa provides exceptional dentistry services from our experienced, skilled, caring, and compassiona...
Channelnews : microsoft leads global takedown of infamous malware tool behind cyber attacksIn a coordinated operation involving Europol, Japan’s JC3, and the US Department of Justice, Microsoft’s Digital Crimes ...
Latests News
Behavioral and neurophysiological aspects of working memory impairment in children with dyslexiaABSTRACT The present study aimed to identify behavioral and neurophysiological correlates of dyslexia which could potent...
Knicks’ og anunoby’s late-game gaffes spoil early heroics in fourthOG Anunoby was enduring another of those quiet offensive games he produces now and again, when the Knicks defensive stop...
Does medicare cover costs of physical therapy sessions?Memorial Day Sale! Join AARP for just $11 per year with a 5-year membership Join now and get a FREE gift. Expires 6/4 G...
Page Unavailable - ABC NewsABC NewsVideoLiveShowsShopLog InStream onOops! Page unavailable.This page either does not exist or is currently unavaila...
Additional sp cid-ci kashmir transferred to acbJAMMU, MAY 28: J&K Home Department on Saturday transferred Additional SP, CID CI Kashmir to J&K Anti Corruption ...