Phosphorus (p) use efficiency in rice is linked to tissue-specific biomass and p allocation patterns
Phosphorus (p) use efficiency in rice is linked to tissue-specific biomass and p allocation patterns"
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT Phosphorus (P) is a non-renewable resource which may be depleted within next few decades; hence high P use efficiency is need of time. Plants have evolved an array of adaptive
mechanisms to enhance external P acquisition and reprioritize internal utilization under P deficiency. Tissue specific biomass and P allocation patterns may affect the P use efficiency in
plants. six rice cultivars were grown in solution culture for 20 days and then were divided into two groups to receive either adequate P or no P that were harvested at 30, 40 and 50 days.
Plants were dissected into various tissues/organs. Two rice cultivars viz Super Basmati (P-inefficient) and PS-2 (P-efficient) were grown in soil with no or 50 mg P kg−1 soil till maturity.
Rice cultivars PS-2 and Basmati-2000 had higher P uptake, utilization efficiency and internal remobilization than other tested cultivars after P omission. Young leaves and roots were the
major sinks while stems and mature leaves were the sources of P during P omission. In conclusion, biomass allocation and P accumulation among various tissues and P remobilization were major
factors responsible for P efficiency. SIMILAR CONTENT BEING VIEWED BY OTHERS EFFECTS OF DIFFERENT PHOSPHORUS AND POTASSIUM SUPPLY ON THE ROOT ARCHITECTURE, PHOSPHORUS AND POTASSIUM UPTAKE,
AND UTILIZATION EFFICIENCY OF HYDROPONIC RICE Article Open access 11 September 2024 GENETIC APPROACHES FOR ASSESSMENT OF PHOSPHORUS USE EFFICIENCY IN GROUNDNUT (_ARACHIS HYPOGAEA_ L.)
Article Open access 13 December 2022 POLYPHOSPHATE FERTILIZER IMPACTS THE ENZYMATIC AND NON-ENZYMATIC ANTIOXIDANT CAPACITY OF WHEAT PLANTS GROWN UNDER SALINITY Article Open access 11 July
2023 INTRODUCTION Phosphorus (P) is the second-most essential element after nitrogen (N) for its impact on the productivity and health of aquatic and terrestrial ecosystems1. Being an
essential element, P is considered as a major driver for optimum crop productivity on arable land around the globe2,3,4. However, suboptimal P availability on arable land forces researchers
to determine the mechanisms for improving P acquisition and utilization, and to exploit these responses for the development of P-efficient cultivars5,6. The identification of desirable crop
plants for adaptation to low-P-input environments may enhance crop productivity and reduce the reliance on costly synthetic P fertilizers6. This approach is especially important for
developing countries with limited resources. Under the current situation, farmers need P-efficient genotypes that perform better than other genotypes with equivalent P inputs7,8,9,10,11.
Many crop plants have evolved morphological, physiological, biochemical and molecular adaptive systems to cope with P-deficiency stress, such as altered root architecture to explore more
soil volume12, and increased carboxylate exudation containing phosphatases, nucleases and various organic acids13,14. These mechanisms and strategies are necessary to liberate or solubilize
Pi from organic and other insoluble pools15, enhance Pi uptake capacity16, recycle internal Pi13,17, remobilize/retranslocate P from mature to young developing organs18,19, and reprioritize
metabolic P utilization20. Cultivars with enhanced P efficiency could be an alternate strategy for overcoming the dilemma of P deficiency. Phosphorus efficiency can be divided into (i) P
acquisition efficiency – the capacity of a cultivar to extract P from soil, and (ii) P utilization efficiency – the capacity of a cultivar to transform the acquired P into biomass/grain
yield21,22. The process of P remobilization from mature leaves frequently occurs during vegetative growth, when there is insufficient soil P available23. However, remobilization can also
happen during the reproductive period, when new sinks are emerging while further P acquisition by plant roots is reduced24. The remobilization process during reproductive growth is generally
associated with foliar senescence to ensure nutrient supply for developing tissues25 and phloem transport is mainly required for the remobilization process, as P is highly mobile in the
phloem26. This mechanism of P remobilization is responsible for inter- and intra-specific differences in crop plants for P utilization. These traits are heritable and can be exploited to
screen plants for high P-efficiency27. Rice (_Oryza sativa_ L.) is a major cereal crop known to provide calories to about one third of the population around the globe. According to
estimates, global rice harvest removes P from fields of worth around $11 billion each year28. Different rice genotypes with higher P-use efficiency can be utilized in rice yield improvement
programs. Enhanced internal P-utilization efficiency is required to complement higher P uptake traits for successful breeding of P efficient rice cultivars29. Screening of existing germplasm
and identification of the responsible P-efficiency mechanism is needed to produce more P-efficient cultivars through conventional as well as molecular breeding ventures. The present study
was conducted to identify P-efficient rice cultivars and evaluate the responsible mechanisms for P efficiency in two independent experiments. RESULTS EXPERIMENT 1 VARIATION IN GROWTH
RESPONSE OF RICE CULTIVARS UNDER P STARVATION The rice cultivars had significant (_P_ ≤ 0.05) variations in plant height, root depth, root dry matter (RDM), shoot dry matter (SDM) and
root:shoot ratio (RSR) under both adequate P (200 µM Pi) and P deficiency (Table 1). Plant height increased linearly with time at both P levels. Averaged across all harvests, PS-2 and
Basmati-2000 had the tallest plants while Super Basmati had the shortest plants at both P levels. Rooting depth of all rice cultivars increased in response to P deficiency relative to
adequate P supply. The cultivar PS-2 had the deepest roots at 39.6 cm and 42.8 cm with adequate P and no P, respectively, and KS-282 had the shallowest rooting depth at 27.4 cm with no P.
The six rice cultivars varied in RDM with Basmati-2000 producing the maximum (0.53 g plant−1) and Super Basmati producing the minimum (0.20 g plant−1) amounts under P deficiency. The SDM of
rice cultivars decreased significantly after exposure to P deficit, but the reduction varied significantly between cultivars. Overall, cultivar PS-2 produced the most SDM (3.31 g plant−1)
under adequate P, while Basmati-2000 produced the most (1.36 g plant−1) with no P supplied. The RSR increased in all tested cultivars under P stress except for KS-282 which had no change at
either P level. The maximum RSR after P omission (0.45) occurred in KSK-434 (Table 1). GENOTYPE-DEPENDENT VARIATION IN BIOMASS PARTITIONING The data for dry matter production of various
plant tissues, i.e., stem, young leaves (tip and base), and mature leaves (tip and base) growing with or without P is presented in Table 2. Interestingly, stem dry matter declined
significantly with P stress in all rice cultivars. With adequate P, rice cultivars PS-2 and Super Basmati produced the maximum (1.49 g plant−1) and minimum (0.97 g plant−1) stem dry matter,
respectively, while the corresponding values under P stress were 0.44 g plant−1 in Basmati-2000 and 0.14 g plant−1 in IR-6. Overall, KSK-434 produced the most biomass in the young leaf tip
(0.48 g plant−1) and young leaf base (0.53 g plant−1) under adequate P supply, while Basmati-2000 produced the maximum biomass (0.20 and 0.25 g plant−1, respectively) after P omission. The
cultivar PS-2 accumulated the most biomass in the mature leaf tip (0.27 g plant−1) and mature leaf base (0.29 g plant−1), and IR-6 produced the least biomass (0.11 g plant−1 and 0.07 g
plant−1, respectively) after P omission. However, under adequate P supply, cultivars KSK-434 and Basmati-2000 produced the maximum (0.40 and 0.44 g plant−1) and minimum (0.25 and 0.28 g
plant−1) biomass in the mature leaf tip and mature leaf base, respectively. Overall, Basmati-2000 produced the most dry matter in young leaves (tip + base), and PS-2 produced the most in
mature leaves (tip + base) under P deficiency. P ACQUISITION AND TISSUE-SPECIFIC PI ALLOCATION The P concentration [P] in different plant tissues, i.e., root, stem, young leaves (tip and
base), and mature leaves (tip and base) at 20, 30, 40 and 50 DAT at each P level is given in Table 3. The [P] in plant roots with no P added increased from 20 to 50 DAT. Overall, PS-2 had
the highest root [P] with 4.47 and 3.13 mg g−1 and IR-6 had the least [P] with 2.40 and 1.66 mg g−1 under adequate and P stress, respectively. Stem [P] decreased with time in all cultivars
under P deficiency. Under P stress, KS-282 had the highest stem [P] (2.73 mg g−1) while Basmati-2000 had the least (1.61 mg g−1). The six rice cultivars varied for [P] in young leaf tips and
basal sections at both P levels. After P omission, PS-2 had the highest [P] with 3.18 mg g−1 in young leaf tips while Basmati-2000 had the least [P] with 2.06 mg g−1. The rice cultivars
KS-282 and Super Basmati had the highest (3.23 mg g−1) and lowest (2.22 mg g−1) [P] in the young leaf base, respectively, among the tested cultivars under P deficiency. The [P] in mature
leaf tips and mature leaf base was higher in IR-6 and KSK-434 (2.36 and 1.73 mg g−1, respectively) after P omission. The cultivars varied significantly (_P_ ≤ 0.05) in P uptake in different
plant parts when grown under adequate P or P stress (Table 4). The highest root P uptake under adequate P or P stress was in PS-2 with 3.77 and 1.38 mg plant−1, respectively. Likewise, PS-2
had higher stem P uptake at adequate P supply and after P stress (6.96 mg plant−1 and 0.73 mg plant−1, respectively). The maximum P uptake (0.63 mg plant−1) by young leaf tips occurred in
PS-2 while the minimum (0.21 mg plant−1) occurred in IR-6 under P stress. Basmati-2000 and Super Basmati had the highest (0.76 mg plant−1) and lowest (0.26 mg plant−1) P uptake by the young
leaf base after P omission. Mature leaf tips accumulated higher P contents than the mature leaf base after P omission. PS-2 had the highest P uptake by the mature tip and basal sections
(0.49 and 0.41 mg plant−1, respectively) during P stress. When comparing P uptake between young and mature leaves, young leaves had higher P uptake at adequate P and under P stress. Averaged
across all harvests, PS-2 had the highest total P uptake (4.10 mg plant−1) while IR-6 had the lowest (1.35 mg plant−1) under P stress. INTERNAL REMOBILIZATION AND UTILIZATION EFFICIENCY OF
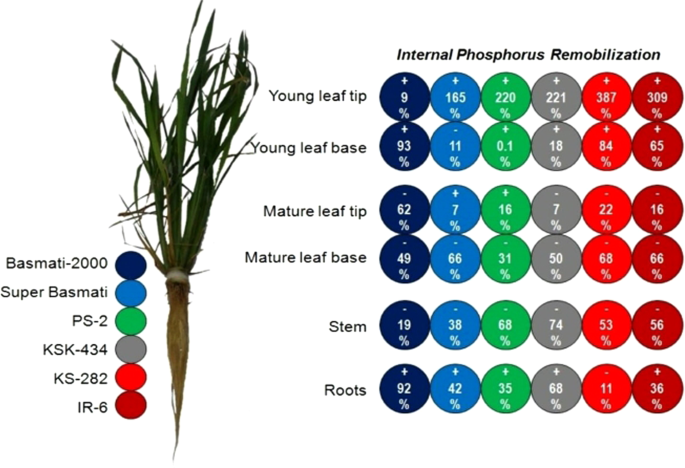
ACQUIRED P BY RICE CULTIVARS The internal P remobilization (IPR) was calculated among various plant tissues at 50 days of transplanting after feeding plants for 20 days with adequate P and
subsequent P omission for next 30 days (Fig. 1). At this time, P had accumulated in the roots and young leaves after remobilization from the stem and mature leaves. Cultivars KSK-434 and
PS-2 remobilized the most stem P reserves (74 and 68%, respectively) after P omission. The lowest IPR from stems occurred in Basmati-2000, but its roots accumulated the most P (92%) during P
omission. Among young leaves, the apical section (tip) accumulated comparatively more P reserves than the basal section in all cultivars except Super Basmati where P contents declined upto
11% for the young leaf base. Nevertheless, basal sections of mature leaves remobilized more P than the mature leaf apical (tip) section in all cultivars except PS-2 and Super Basmati where P
accumulation (16 and 7%, respectively) occurred in the mature leaf tip. Phosphorus utilization efficiency (PUTE) varied from 12.1% (Super Basmati) to 21.7% (PS-2) with a mean value of 16.9%
(Fig. 2). EXPERIMENT 2 Two rice cultivars contrasting in P utilization efficiency - Super Basmati (P-inefficient) and PS-2 (P-efficient) - were selected from Experiment 1 and grown at two P
levels (0 and 50 mg P kg−1) in soil-filled pots to maturity to investigate their response for paddy production, P acquisition and utilization efficiencies. YIELD AND RELATED ATTRIBUTES
Phosphorus deficiency significantly (_P_ ≤ 0.05) reduced rice yield and related attributes, i.e., plant height, tiller number and panicle length, in both rice cultivars (Table 5, Fig. 3).
However, the magnitude of reduction was higher in cultivar Super Basmati than cultivar PS-2. Plant height of Super Basmati and PS-2 varied from 42.3–48.0 cm and 63.0–73.0 cm under deficient
and adequate P supply, respectively. Super Basmati had more tillers under P deficiency than PS-2 (13.7 vs. 9.3). The panicle length of Super Basmati and PS-2 increased from 8.7 to 14.3 cm
and 13.7 to 18.0 cm in response to deficient and adequate P, respectively. PS-2 produced the highest paddy yield (18.8 vs. 29.0 g pot−1) as compared to Super Basmati (8.7 vs. 15.8 g pot−1)
in unfertilized and P-fertilized plants, respectively. Straw yield in PS-2 and Super Basmati declined in unfertilized plants (38.2 and 20.5 g pot−1) and increased in response to P
fertilization (44.4 and 30.9 g pot−1), respectively. The biological yield (straw + paddy) of PS-2 and Super Basmati was 73.4 and 46.7 g pot−1 in P-fertilized plants and 57.0 and 29.2 g pot−1
in P-deficient plants, respectively. PHOSPHORUS UPTAKE AND EFFICIENCY RELATIONS OF RICE CULTIVARS Phosphorus uptake in paddy, straw, and total (paddy + straw) by rice cultivars grown under
soil conditions with deficient and adequate P supply is depicted in Fig. 3. The rice cultivars and P levels had significant (_P_ ≤ 0.05) main and interactive effects on P uptake by paddy and
straw (Table 5). Average P uptake by both straw and paddy was about two-fold higher in plants with adequate P supply than deficient levels. The paddy P uptake in PS-2 and Super Basmati was
130.81 and 60.27 mg pot−1 under adequate P and 80.87 and 27.25 mg pot−1 under P deficiency, respectively. Under adequate P, straw P uptake in PS-2 and Super Basmati was 95.50 and 61.54 mg
pot−1 and 59.44 and 19.59 mg pot−1 under P deficiency, respectively. The values for total P uptake ranged from 140.31 to 226.30 mg pot−1 for PS-2 and 46.84 to 121.81 mg pot−1 for Super
Basmati under deficient and adequate P, respectively. Both rice cultivars responded differently for P-efficiency relations – P acquisition efficiency (PACE), P utilization efficiency (PUTE)
and P stress factor (PSF) – at the two soil P levels (Fig. 4). The rice cultivar PS-2 had higher values than Super Basmati for PACE (14.3 vs. 9.4%) and PUTE (64.7 vs. 55.8%). The PSF
describes the percent reduction in paddy production in response to P deficiency in the rooting medium. It distinguishes between P-responsive and non-responsive genotypes and elucidates the
capacity of a genotype to produce biomass with P addition. Super Basmati had a larger PSF (44.2%) than PS-2 (35.3%). DISCUSSION Limited P availability to plants on arable land is a major
nutritional constraint hindering plant growth and crop yields. Significant variation for P absorption and utilization among crop species and even genotypes within the same species is well
documented4,5,10,30,31. The selection/identification of cultivars that can absorb and use P efficiently is a promising strategy to cope with environments deficient in bio-available P12.
Phosphorus is a highly mobile element within plants that is readily remobilized/re-translocated from metabolically inactive to active sites to sustain plant growth under P deficiency32. In
this study, we investigated the hypothesis that variation in P acquisition and remobilization is due to adaptive mechanisms that cause differential P utilization in rice cultivars. In
Experiment 1, root and shoot dry matter (DM) production under P deficiency varied between rice cultivars. Dry matter production under low-P supply is an excellent parameter for screening
germplasm for higher yields and P efficiency8,33. Two efficient cultivars, PS-2 and Basmati-2000, produced more DM than Super Basmati and IR-6 under P starvation, which can be attributed to
an increased rate of P remobilization to growing sites, i.e., young leaves and roots. Remobilization of stored P in the stem and older leaves to metabolically active sites may supplement the
restricted P supply under P deficiency13,34,35. This was confirmed at all harvests by the differential P redistribution and accumulation in various plant tissues during P omission (Fig. 1,
Table 4). Variations in shoot biomass under varying levels of P among different rice cultivars have been reported36. Roots are critical for plant growth and are directly exposed to the soil
environment. As P mobility in soil is limited, higher plant root growth and changes in root morphology is helpful for more P uptake. Rice cultivar PS-2 had the deepest roots and the most
root dry matter when exposed to P deficiency (Table 1). Under P starvation, root length and root hair density significantly increased37 for better P absorption. The findings of Bates and
Lynch38 suggested that increased root growth is associated with improved plant performance under low P by exploring a larger volume of soil. The root:shoot ratio (RSR) is a further
indication of the partitioned plant growth into roots and shoots to differential P supply. The RSR increased in all tested cultivars under P stress while KSK-434 had the maximum RSR.
Generally, under low-P supply, plant growth is suppressed. However, the relative effect is most dominant in the case of shoot growth. Consequently, RSR increases significantly in low-P
environments39 and is an excellent index for partitioning photosynthesized carbon between above – and below – ground plant parts. Root density and RSR generally increased under P deficiency,
thus favoring P acquisition by plants13,40. Plants often allocate a greater portion of their biomass to the roots in response to P deficient environment. Such acclamatory response is a
consequence of metabolic changes in shoot and an adjustment of carbohydrate transport to the root. P deficiency alter root-to-shoot biomass ratio by accumulation of carbohydrates in leaves
and roots41. These findings were confirmed in the current study. Increased remobilization of absorbed P among various tissues within plants under P deficiency might be a mechanism for better
P efficiency in cultivars differing in P acquisition and utilization32,42. In Experiment 1, plants were grown for 20 days with adequate P (200 µM Pi), after which P was omitted from the
solution for the next 30 days. As plants were unable to uptake more P from the medium, they had to remobilize previously acquired P from various plant tissues. Molecular mechanisms involved
in mobilizing P from different plant tissues/organs are well reported15,43,44,45. Organic pools of P in plant tissues mainly exist as nucleic acids, phospholipids, phosphorylated proteins
and P-ester metabolites15. Under P deficiency, phospholipids have greater potential to serve as an alternate source of P which remobilizes towards young growing plant tissues46. This
phenomenon of replacing phospholipids in cell membranes with sulfolipids and galactolipids is known as membrane-lipid remodeling43. Lipid remodeling occurs both in roots and shoots of
P-efficient rice genotypes under low-P conditions resulting in enhanced PUE44. Ribonucleases and intracellular phosphatases had crucial role in determining the amount of P to be remobilized
from older to young green leaves and developing grains45. During P omission, P accumulated in the roots and young leaves after remobilization from the stem and mature leaves (Fig. 1, Table
4). Cultivars KSK-434 and PS-2 remobilized the most stem P reserves (74 and 68%, respectively) after P omission. Within young leaves, the apical section (tip) accumulated comparatively more
P than the basal section (Table 4). Based on differential P contents in various plant tissues during P omission; it could be assumed that P remobilization from the metabolically inactive
pool to the active pool was initially more intensive in mature leaves, stem, roots, and young leaves. With persistent P stress, the stems recorded the maximum reduction in P contents. An
increase or even maintenance of root P content during P stress may be related to greater allocation of assimilates to roots47. The higher P contents in roots and young leaves of rice under
adequate P illustrates that young tissues have better storage capacity than older tissues17. Moreover, enhanced metabolic activities of young tissues make them stronger sinks for the already
absorbed P. After removing P from the nutrient solution, plant growth may occur due to remobilization and utilization of absorbed P48. According to Martinez _et al_.49, young leaves and
roots were the major sinks of stored P in soybean cultivars. Under moderate P deficiency, the transfer of P increased in the shoots of potato plants, while severely stressed plants retained
P in the roots50. Although, more P acquisition form P-deficient medium is necessary for tolerance of a genotype to low P, however efficient remobilization of acquired P to different growing
tissues can make a genotype well adapted to low-P environment51. Efficient internal utilization of P is generally attributed because of high photosynthetic activity per unit of P and more
efficient P remobilization from older to young leaves52. Moreover, higher remobilization of P from older leaves to young green leaves in P-efficient rice genotypes is important to produce
more photosynthates which ultimately contribute for biomass/yield production53. In Experiment 2, two rice cultivars selected from the hydroponic study – Super Basmati (P-inefficient) and
PS-2 (P-efficient) – were grown in soil to maturity to investigate their response for paddy production, P acquisition, and utilization efficiencies. It was clear from the results that the
high-yielding cultivar PS-2 had more paddy P uptake and the low-yielding cultivar Super Basmati had the lowest paddy P uptake under both P levels. Fageria54 also reported increased P uptake
in grains of high-yielding rice genotypes at low P. According to Baligar _et al_.55, more P uptake in plants grown at high P compared with those at low P is due to an increased root fine
hair density. The rice cultivars varied in P-efficiency relations (PACE, PUTE, and PSF) under P stress. The high-yielding PS-2 had greater PACE and PUTE than the low-yielding Super Basmati.
Phosphorus utilization and uptake efficiencies are important for selecting genotypes under P-deficiency stress. The P-use efficiency of cultivars is associated with the amount of dry matter
produced per unit of absorbed P56. The variation between cultivars for PUE might be due to differential P absorption characteristics or distribution patterns within plants. Both cultivars
varied in their relative reduction in paddy yield due to P deficiency (PSF). The PSF value distinguishes between P-responsive and non-responsive cultivars and the comparative ability of a
cultivar to produce biomass with P addition4,5,13. Cultivar PS-2 had a lower PSF value than Super Basmati under P stress (35% vs. 44%). Under P deficiency, PS-2 produced more paddy yield
than Super Basmati and responded well to P application. Super Basmati was less desirable due to poor performance under varying P supply. PS-2 also had higher P uptake than Super Basmati,
indicating an important contributing trait to its high P-efficiency. The lower P-efficiency of Super Basmati was further supported by its higher PSF value than PS-2. Fageria _et al_.30
reported variation in the abilities of lowland rice cultivars to absorb P, and its remobilization and efficient utilization overtime. Plant traits associated with efficient P acquisition and
internal utilization are heritable and can be exploited more extensively to develop P-efficient cultivars27. CONCLUSIONS Significant genetic variation exists among the tested rice cultivars
for P efficiency. Rice cultivars with higher P uptake, acquisition efficiency, utilization efficiency and lower PSF values were efficient (low-P tolerant) and thus more desirable for
adaptation to soils low in available P. The cultivars with high biomass/paddy yield at both P levels (PS-2 and Basmati-2000) have better internal remobilization of absorbed P and
photosynthate translocation to young/active plant tissues. Hence, P partitioning and biomass allocation to roots should be considered when screening germplasm in P-efficiency programs.
MATERIALS AND METHODS EXPERIMENT 1 PLANT MATERIAL, GROWTH CONDITIONS, AND MANAGEMENT Seeds of six rice cultivars (Basmati-2000, Super Basmati, PS-2, KSK-434, KS-282 and IR-6) were kindly
provided by the Rice Research Institute, Kala Shah Kaku, Pakistan. Seeds were surface sterilized with 3% sodium hypochlorite solution and sown in polyethylene foil-lined metal trays
containing two inches of washed riverbed sand. Distilled water was used to maintain optimum moisture for seed germination and seedling establishment. Two weeks after seed germination, the
root systems of seedlings were carefully rinsed in distilled water to remove any adhering sand. Uniform seedlings were transferred to 25 L plastic tubs containing modified Johnson’s nutrient
solution in a completely randomized design. The composition of the full-strength nutrient solution (pH 5.5) was 5.0 mM N, 3.5 mM K, 1.5 mM Ca, 0.5 mM Mg, 2.05 mM S, 50 µM Cl, 0.5 µM Mo,
25.0 µM B, 0.5 µM Cu, 2.0 µM Mn, 2.0 µM Zn, and 50.0 µM Fe as Fe-EDTA. The seedlings were held with foam plugs in the holes of a thermopore sheet placed at the top of each tub. Initially,
plants grew for 20 days after transplanting (DAT) with adequate P supply (200 µM Pi). At 20 DAT, three replications of each cultivar were harvested, and the remaining plants were divided
into two groups that received nutrient solution with either 200 µM Pi or without Pi and harvested at 30, 40 and 50 DAT. At each harvest, plant roots were rinsed (2–3 times) with distilled
water to remove the nutrient solution. Shoot samples were washed with distilled water, blotted dry, and then separated into the stem, young (top) leaves and mature (lower) leaves. Young and
mature leaves were further dissected from the middle into leaf tip (upper half) and leaf base (lower half). Plant material was dried at 70 °C for 48 h in a forced air-driven oven and stored
under desiccation until weighing for dry biomass. EXPERIMENT 2 EXPERIMENTAL SITE, DESIGN AND CROP MANAGEMENT Two rice cultivars contrasting in P utilization efficiency were selected from
Experiment 1 [Super Basmati (P-inefficient) and PS-2 (P-efficient)] and grown in plastic pots containing 7 kg of soil. Bulk soil from the top 15 cm was collected from the research area of
the Institute of Soil and Environmental Sciences, University of Agriculture, Faisalabad, Pakistan. A composite sample of collected soil was subjected to various physicochemical properties.
In brief, the soil was a clay loam, non-saline (ECe, 2.12 dS m−1), alkaline in soil reaction (pHs, 7.76), low in Kjeldahl nitrogen (0.04%), organic matter (0.71%) and available phosphorus
(2.01 mg kg−1) and high in available potassium (160 mg kg−1). The experiment was arranged in a completely randomized factorial design having three replicates. All pots were irrigated with
canal water, applied from the top to soil saturation and kept for two days for soil settling. The required amounts of P and K were added at the time of pot filling while N was applied in
three equal splits. Both rice cultivars were grown at two P levels [i.e., adequate (P addition at 50 mg kg−1 soil) and deficient (native soil P at 5.30 mg kg−1 without external P addition)].
Seeds of rice cultivars were sown in nursery trays containing similar soil to the pots. At the three-leaf stage, three uniform and healthy seedlings of each cultivar were transplanted to
each pot. A water layer (1–2 cm) on the soil surface was maintained during the entire crop period. At maturity, plants were harvested and threshed manually to separate paddy from straw.
After recording yield and related attributes, plant material was oven dried at 70 °C till further analysis. SOIL ANALYSIS In order to determine physicochemical properties of soil used in
experiment 2, soil sample was air-dried and grounded to pass through a 2 mm sieve. Soil texture was determined by performing mechanical analysis of soil separates (sand, silt, and clay)
using hydrometer method by dispersing soil in sodium hexametaphosphate solution57. Soil reaction of the saturated soil paste (pHs) and electrical conductivity of saturation extract (ECe)
were determined with pH and EC meter, respectively58. Soil available phosphorus and potassium were estimated using ammonium bicarbonate-diethylene triamine penta-acetic acid (AB-DTPA) as
extracting solution59. Kjeldahl nitrogen was determined following the method described by Jackson60. Soil organic matter (SOM) was quantified by chromic acid digestion according to
Walkley-Black method61. PLANT ANALYSIS Oven-dried plant material from both experiments was finely ground in a sample grinder (IKA Werke, Wilmington, USA). Ground samples (0.3 g each) were
wet digested using 10 mL of di-acid digestion mixture [(HNO3:HClO4 (5:1, _v/v_)]. Total P concentration in plant samples was determined following the vanadate–molybdate method62 using
UV-visible spectrophotometer (Shimadzu UV-VIS 1201, Shimadzu Co. Kyoto, Japan). CALCULATION METHODS The following P-efficiency relations were calculated to establish the relationship between
P levels and plant growth of rice cultivars, i.e., P uptake63, internal P remobilization64, acquisition efficiency2, utilization efficiency65, and stress factor66 using the following
formulas: * Phosphorus uptake (PU) $${\rm{PU}}({\rm{mg}}\,{{\rm{plant}}}^{-1})={\rm{P}}\,{\rm{concentration}}({\rm{mg}}\,{{\rm{g}}}^{-1})\times
{\rm{dry}}\,\text{matter}(g\,{{\rm{plant}}}^{-1})$$ * Internal phosphorus remobilization (IPR) $${\rm{IPR}}\,( \%
)=\frac{{\rm{P}}\,{{\rm{uptake}}}_{{\rm{before}}{\rm{P}}{\rm{omission}}}-{\rm{P}}\,{{\rm{uptake}}}_{{\rm{after}}{\rm{P}}{\rm{omission}}}}{{\rm{P}}\,{{\rm{uptake}}}_{{\rm{before}}{\rm{P}}{\rm{omission}}}}\times
100$$ * Phosphorus acquisition efficiency (PACE) $${\rm{PACE}}( \%
)=\frac{{\rm{P}}\,{{\rm{uptake}}}_{{\rm{adequate}}{\rm{P}}}-{\rm{P}}\,{{\rm{uptake}}}_{{\rm{deficient}}{\rm{P}}}}{{\rm{Quantity}}\,{\rm{of}}\,{\rm{P}}\,{\rm{applied}}}\times 100$$ *
Phosphorus utilization efficiency (PUTE) $${\rm{PUTE}}( \% )=\frac{{\rm{Paddy}}\,{{\rm{yield}}}_{{\rm{deficient}}{\rm{P}}}}{{\rm{Paddy}}\,{{\rm{yield}}}_{{\rm{adequate}}{\rm{P}}}}\times
100$$ * Phosphorus stress factor (PSF) $${\rm{PSF}}( \%
)=\frac{{\rm{Paddy}}\,{{\rm{yield}}}_{{\rm{adequate}}{\rm{P}}}-{\rm{Paddy}}\,{{\rm{yield}}}_{{\rm{deficient}}{\rm{P}}}}{{\rm{Paddy}}\,{{\rm{yield}}}_{{\rm{adequate}}{\rm{P}}}}\times 100$$
STATISTICAL ANALYSIS The computer software STATISTIX 8.1 (Analytical Software, Inc., Tallahassee, FL, USA) was used to perform statistical analysis following the methods of Steel _et al_.67.
All data reported in this study are the means of three replicates and presented with standard errors. First experiment was conducted following completely randomized desing and the
significant differences among rice cultivars regarding growth, biomass production and P accumulation in various plant tissues were differentiated at each P level using one-way analysis of
variance (ANOVA) technique with an associated least significant difference test at 5% probability level (LSD, _P_ ≤ 0.05). In second experiment, completely randomized design with factorial
arrangement was employed and the results for yield, P uptake and P-efficiency relations were compared by two-way ANOVA technique with an associated LSD test at _P_ ≤ 0.05. The graphical
presentation of data was performed usign Microsoft Office (Redmond, WA, USA). DATA AVAILABILITY All data generated or analysed during this study are included in this published article.
REFERENCES * Brady, N. C. & Weil, R. R. _The Nature and Properties of Soils_ (14th Ed). Prentice Hall, Upper Saddle River, New Jersey (USA 2008). * Baligar, V., Fageria, N. & He, Z.
Nutrient use efficiency in plants. _Comm. Soil Sci. Plant Analysis._ 32, 921–950 (2001). Article CAS Google Scholar * Stewart, W. M., Dibb, D. W., Johnston, A. E. & Smyth, T. J. The
contribution of commercial fertilizer nutrients to food production. _Agron. J._ 97, 1–6 (2005). Article Google Scholar * Irfan, M., Shah, J. A. & Abbas, M. Evaluating the performance
of mungbean genotypes for grain yield, phosphorus accumulation and utilization efficiency. _J. Plant Nutr._ 40, 2709–2720 (2017). Article CAS Google Scholar * Abbas, M., Irfan, M., Shah,
J. A. & Memon, M. Y. Intra-specific variations among wheat genotypes for phosphorus use efficiency. _Asian J. Agric. Biol._ 6, 35–45 (2018). Google Scholar * Irfan, M., Abbas, M., Shah,
J. A. & Memon, M. Y. Internal and external phosphorus requirements for optimum grain yield are associated with P-utilization efficiency of wheat cultivars. _Commun. Soil Sci. Plant
Anal._ 49, 2843–2853 (2018). Article CAS Google Scholar * Dawson, C. J. & Hilton, J. Fertiliser availability in a resource-limited world: production and recycling of nitrogen and
phosphorus. _Food Pol._ 36, S14–S22 (2011). Article Google Scholar * Abbas, M., Irfan, M. & Shah, J. A. Differential performance of wheat genotypes for grain yield, phosphorus uptake
and utilization at low and high phosphorus levels. _Proc. Pak. Acad. Sci.: B. Life Environ. Sci._ 55, 55–64 (2018). Google Scholar * Ozturk, L., Eker, S., Torun, B. & Cakmak, I.
Variation in phosphorus efficiency among 73 bread and durum wheat genotypes grown in a phosphorus-deficient soil. _Agron. J._ 74, 562–564 (2005). Google Scholar * Aziz, T., Finnegan, P. M.,
Lambers, H. & Jost, R. Organ‐specific phosphorus‐allocation patterns and transcript profiles linked to phosphorus efficiency in two contrasting wheat genotypes. _Plant, Cell Environ._
37, 943–960 (2014). Article CAS Google Scholar * Irfan, M. _et al_. Differential performance of lowland rice cultivars for phosphorus uptake and utilization efficiency under hydroponic
and soil conditions. _Intr. J. Agric. Biol._ 21, 703–710 (2019). CAS Google Scholar * Aziz, T., Steffens, D., Rahmatullah & Schubert, S. Variation in phosphorus efficiency among
_Brassica_ cultivars II: Changes in root morphology and carboxylate exudation. _J. Plant Nutr._ 34, 2127–2138 (2011). Article CAS Google Scholar * Aziz, T., Ahmed, I., Farooq, M.,
Maqsood, M. A. & Sabir, M. Variation in phosphorus efficiency among _Brassica_ cultivars I: Internal utilization and phosphorus remobilization. _J. Plant Nutr._ 34, 2006–2017 (2011).
Article CAS Google Scholar * Pearse, S. J., Veneklaas, E. J., Cawthray, G. R., Bolland, M. D. A. & Lambers, H. Carboxylate release of wheat, canola and 11 grain legume species as
affected by phosphorus status. _Plant Soil._ 288, 127–139 (2006). Article CAS Google Scholar * Veneklaas, E. J. _et al_. Opportunities for improving phosphorus-use efficiency in crop
plants. _New Phytol._ 195, 306–320 (2012). Article CAS PubMed Google Scholar * Shen, J., Li, H., Neumann, G. & Zhang, F. Nutrient uptake, cluster root formation and exudation of
protons and citrate in _Lupinus albus_ as affected by localized supply of phosphorus in a split-root system. _Plant Sci._ 168, 837–845 (2005). Article CAS Google Scholar * Abel, S.,
Ticconi, C. A. & Delatorre, C. A. Phosphate sensing in higher plants. _Physiol. Plant._ 115, 1–8 (2002). Article CAS PubMed Google Scholar * Abbas, M., Shah, J. A., Irfan, M. &
Memon, M. Y. Remobilization and utilization of phosphorus in wheat cultivars under induced phosphorus deficiency. _J. Plant Nutr._ 41, 1522–1533 (2018). Article CAS Google Scholar *
Irfan, M. _et al_. Biomass and phosphorus accumulation, partitioning and remobilization during grain development in wheat under phosphorus deficiency. _Intr. J. Agric. Biol._ 21, 351–358
(2019). CAS Google Scholar * Aziz, T., Lambers, H., Nicol, D. & Ryan, M. H. Mechanisms for tolerance of very high tissue phosphorus concentrations in _Ptilotus polystachyus_. _Plant,
Cell Environ._ 38, 790–799 (2015). Article CAS Google Scholar * Vance, C. P., Uhde‐Stone, C. & Allan, D. L. Phosphorus acquisition and use: critical adaptations by plants for securing
a nonrenewable resource. _New Phytol._ 157, 423–447 (2003). Article CAS Google Scholar * Manschadi, A. M., Kaul, H. P., Vollmann, J., Eitzinger, J. & Wenzel, W. Developing
phosphorus-efficient crop varieties: An interdisciplinary research framework. _Field Crops Res._ 162, 87–98 (2014). Article Google Scholar * Abdallah, M. _et al_. Effect of mineral sulphur
availability on nitrogen and sulphur uptake and remobilization during the vegetative growth of _Brassica napus_ L. _J. Exp. Bot._ 61, 2635–2646 (2010). Article CAS PubMed PubMed Central
Google Scholar * Malagoli, P., Laine, P., Rossato, L. & Ourry, A. Dynamics of nitrogen uptake and mobilization in field-grown winter oil seed rape (_Brassica napus_ L.) from stem
extension to harvest: I. Global N flows between vegetative and reproductive tissues in relation to leaf fall and their residual N. _Annals Bot._ 95, 853–861 (2005). Article CAS Google
Scholar * Avice, J.-C. & Etienne, P. Leaf senescence and nitrogen remobilization efficiency in oil seed rape (_Brassica napus_ L.). _J. Exp. Bot._ 65, 3813–3824 (2014). Article PubMed
Google Scholar * White, P. J. Long-distance transport in the xylem and phloem. In: Marschner, P. (ed) Marschner’s Mineral Nutrition of Higher Plants (3rd ed.) Elsevier, Berlin, pp. 49–70
(2012). * Yaseen, M. & Malhi, S. S. Variation in yield, phosphorus uptake, and physiological efficiency of wheat genotypes at adequate and stress phosphorus levels in soil. _Commun. Soil
Sci. Plant Anal._ 40, 3104–3120 (2009). Article CAS Google Scholar * Rose, T. J., Pariasca-Tanaka, J., Rose, M. T., Fukuta, Y. & Wissuwa, M. Genotypic variation in grain phosphorus
concentration, and opportunities to improve P-use efficiency in rice. _Field Crops Res._ 119, 154–160 (2010). Article Google Scholar * Vellaikumar, S. & Malarvizhi, P. Phosphorus use
efficiency of selected rice varieties. _Trends Biosci._ 10, 6662–6677 (2017). Google Scholar * Fageria, N. K., Santos, A. B. & Heinemann, A. B. Lowland rice genotypes evaluation for
phosphorus use efficiency in tropical lowland. _J. Plant Nutr._ 34, 1087–1095 (2011). Article CAS Google Scholar * Akhtar, M. S., Oki, Y., Nakashima, Y., Adachi, T. & Nishigaki, M.
Phosphorus stress-induced differential growth, and phosphorus acquisition and use efficiency by spring wheat cultivars. _Commun. Soil Sci. Plant Analysis._ 47, 15–27 (2016). Article CAS
Google Scholar * Akhtar, M. S., Oki, Y. & Adachi, T. Intra-specific variations of phosphorus absorption and remobilization, P forms, and their internal buffering in _Brassica_ cultivars
exposed to a P‐stressed environment. _J. Integ. Plant Biol._ 50, 703–716 (2008). Article CAS Google Scholar * Rengel, Z. & Marschner, P. Nutrient availability and management in the
rhizosphere: exploiting genotypic differences. _New Phytol._ 168, 305–312 (2005). Article CAS PubMed Google Scholar * Marschner, P. _Mineral nutrition of higher plants_. (2nd ed.).
Academic Press, (London 1995). * Peng, Z. & Li, C. Transport and partitioning of phosphorus in wheat as affected by P withdrawal during flag-leaf expansion. _Plant Soil._ 268, 1–11
(2005). Article CAS Google Scholar * Gill, M. A., Mansoor, S. & Aziz, T. Rahmatullah & Akhtar, M. S. Differential growth response and phosphorus utilization efficiency of rice
genotypes. _Pak. J. Agric. Sci._ 39, 83–87 (2002). Google Scholar * Ma, Z., Bielenberg, D. G., Brown, K. M. & Lynch, J. P. Regulation of root hair density by phosphorus availability in
_Arabidopsis thaliana_. _Plant, Cell Environ_. 24, 459–467 (2201). * Bates, T. R. & Lynch, J. P. Root hairs confer a competitive advantage under low P availability. _Plant Soil._ 236,
243–250 (2001). Article CAS Google Scholar * Lu, Y., Wassmann, R., Neub, H. U. & Breno, C. S. Impact of phosphorus supply on root exudation, aerenchyma formation, and methane emission
of rice plants. _Biogeochem._ 47, 203–218 (1999). Google Scholar * Fohse, D., Claassen, N. & Jungk, A. Phosphorus efficiency of plants. II. Significance of root radius, root hairs and
cation anion balance for phosphorus influx in seven plant species. _Plant Soil._ 123, 261–272 (1991). Article Google Scholar * Hermans, C., Hammond, J. P., White, P. J. & Verbruggen,
N. How do plants respond to nutrient shortage by biomass allocation? _Trends Plant Sci._ 11, 610–617 (2006). Article CAS PubMed Google Scholar * Lambers, H., Brundrett, M. C., Raven, J.
A. & Hopper, S. D. Plant mineral nutrition in ancient landscapes: High plant species diversity on infertile soils is linked to functional diversity for nutritional strategies. _Plant
Soil._ 334, 11–31 (2010). Article CAS Google Scholar * Tjellstrom, H., Andersson, M. X., Larsson, K. E. & Sandelius, A. S. Membrane phospholipids as a phosphate reserve: the dynamic
nature of phospholipid-to-digalactosyl diacylglycerol exchange in higher plants. _Plant Cell Environ._ 31, 1388–1398 (2008). Article CAS PubMed Google Scholar * Mehra, P., Pandey, B. K.
& Giri, J. Comparative morphophysiological analyses and molecular profiling reveal Pi-efficient strategies of a traditional rice genotype. _Front. Plant Sci._ 6, 1184 (2016). Article
PubMed PubMed Central Google Scholar * Jeong, K. _et al_. Phosphorus remobilization from rice flag leaves during grain filling: an RNA-seq study. _Plant Biotechnol. J._ 15, 15–26 (2017).
Article CAS PubMed Google Scholar * Zhang, Y. _et al_. Silicon compensates phosphorus deficit-induced growth inhibition by improving photosynthetic capacity, antioxidant potential, and
nutrient homeostasis in tomato. _Agronomy._ 9, 733 (2019). Article Google Scholar * Burauel, P., Wieneke, J. & Ffihr, F. Carbohydrate status in roots of two soybean varieties: a
possible parameter to explain different efficiencies concerning phosphate uptake. _Plant Soil._ 123, 169–174 (1989). Article Google Scholar * Rausch, C. & Bucher, M. Molecular
mechanisms of phosphate transport in plants. _Planta_ 216, 23–37 (2002). Article CAS PubMed Google Scholar * Martinez, H. E. P., Novais, R. F., Rodrigues, L. A. & do Sacramento, L.
V. S. Phosphate forms in plant and their internal buffering in five soybean cultivars. _Revista Brasil. Ciencia Solo._ 29, 249–257 (2005). Article CAS Google Scholar * Cogliati, D. H.
& Clarkson, D. T. Physiological changes and phosphate uptake by potato plants during development of and recovery from phosphate deficiency. _Physiol. Plant._ 58, 287–294 (1983). Article
Google Scholar * Wu, P., Shou, H., Xu, G. & Lian, X. Improvement of phosphorus efficiency in rice on the basis of understanding phosphate signaling and homeostasis. _Curr Opin. Plant
Biol._ 16, 205–212 (2013). Article CAS PubMed Google Scholar * Stigter, K. A. & Plaxton, W. C. Molecular mechanisms of phosphorus metabolism and transport during leaf senescence.
_Plants_ 4, 773–798 (2015). Article CAS PubMed PubMed Central Google Scholar * Assuero, S. G., Mollier, A. & Pellerin, S. The decrease in growth of phosphorus-deficient maize leaves
is related to a lower cell production. _Plant Cell Environ._ 27, 887–895 (2004). Article CAS Google Scholar * Fageria, N. K. Yield and yield components and phosphorus use efficiency of
lowland rice genotypes. _J. Plant Nutr._ 37, 979–989 (2014). Article CAS Google Scholar * Baligar, V. C., Fageria, N. K. & Elrashidi, M. Toxicity and nutrient constraints on root
growth. _Hort. Sci._ 33, 960–965 (1998). Article CAS Google Scholar * Siddiqi, M. Y. & Glass, A. D. M. Utilization index: A modified approach to the estimation and comparison of
nutrient utilization efficiency in plants. _J. Plant Nutr._ 4, 289–302 (1981). Article Google Scholar * Bouyoucos, G. J. Hydrometer method improved for making particle size analysis of
soils. _Agron. J._ 54, 464–465 (1962). Article Google Scholar * Anderson, J. M. & Ingram, J. A. I. _Tropical soil biology and fertility_. Wallingford, UK: CAB International (1993). *
Soltanpour, P. N. & Workman, S. Modification of the NaHCO3 DTPA soil test to omit carbon black. _Commun. Soil Sci. Plant Anal._ 10, 1411–1420 (1979). Article CAS Google Scholar *
Jackson, M. L. _Soil chemical analysis_. p. 151–153. Englewood Cliffs, NJ: Prentice Hall Inc. (1962). * Nelson, D. W. & Sommers, L. E. Total carbon, organic carbon and organic matter. In
_Methods of soil analysis_, Part 2, eds. Page, A. L., Miller, R. H. & Keeney, D. R. p. 539–579, Chemical and microbiological properties. Madison, Wisconsin, American Society of Agronomy
(USA 1982). * Chapman, H. D. & Pratt, P. F. _Methods of analysis for soils, plantsand waters_. University of California, Division of Agricultural Science, Riverside, (USA 1961). *
Zhang, Z. _et al_. Differential responses of conventional and Bt-transgenic cotton to potassium deficiency. _J. Plant Nutr._ 30, 659–670 (2007). Article CAS Google Scholar * Maillard, A.
_et al_. Leaf mineral nutrient remobilization during leaf senescence and modulation by nutrient deficiency. _Front. Plant Sci._ 6, 317 (2015). Article PubMed PubMed Central Google Scholar
* Gunes, A., Inal, A., Alpaslan, M., Cakmak, I. & Walton, P. E. G. Genotypic variation in phosphorus efficiency between wheat cultivars grown under greenhouse and field conditions.
_Soil Sci. Plant Nutr._ 52, 470–478 (2006). Article CAS Google Scholar * Hunt, R. _Plant growth analysis_. Edwards Arnold Publ. Ltd., (London 1978). * Steel, R. G. D., Torrie, J. H. &
Dicky, D. A. _Principles and Procedures of Statistics-A Biometrical Approach_. McGraw-Hill Book Inter. Co., (Singapore 1997). Download references AUTHOR INFORMATION AUTHORS AND AFFILIATIONS
* Institute of Soil and Environmental Sciences, University of Agriculture, Faisalabad, 38040, Pakistan Muhammad Irfan, Tariq Aziz, Muhammad Aamer Maqsood & Hafiz Muhammad Bilal * Soil
and Environmental Sciences Division, Nuclear Institute of Agriculture, Tandojam, 70060, Pakistan Muhammad Irfan * The UWA Institute of Agriculture and UWA School of Agriculture and
Environment, The University of Western Australia, Perth, WA, 6001, Australia Tariq Aziz & Kadambot H. M. Siddique * Department of Environmental Sciences, University of Okara, Okara,
56300, Pakistan Hafiz Muhammad Bilal * National Engineering Laboratory for Improving Quality of Arable Land, Institute of Agri. Resources and Regional Planning, Chinese Academy of
Agricultural Sciences, Beijing, 100081, China Minggang Xu Authors * Muhammad Irfan View author publications You can also search for this author inPubMed Google Scholar * Tariq Aziz View
author publications You can also search for this author inPubMed Google Scholar * Muhammad Aamer Maqsood View author publications You can also search for this author inPubMed Google Scholar
* Hafiz Muhammad Bilal View author publications You can also search for this author inPubMed Google Scholar * Kadambot H. M. Siddique View author publications You can also search for this
author inPubMed Google Scholar * Minggang Xu View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS M.I., T.A. and M.A.M. planed the experiment,
M.I. and H.M.B. conducted the experiment and analyze data, M.I. and T.A. wrote the paper and K.S. and M.X. helps in data analysis and paper writing. CORRESPONDING AUTHORS Correspondence to
Tariq Aziz or Minggang Xu. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral with
regard to jurisdictional claims in published maps and institutional affiliations. RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons Attribution 4.0
International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the
source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative
Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by
statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit
http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Irfan, M., Aziz, T., Maqsood, M.A. _et al._ Phosphorus (P) use efficiency in rice
is linked to tissue-specific biomass and P allocation patterns. _Sci Rep_ 10, 4278 (2020). https://doi.org/10.1038/s41598-020-61147-3 Download citation * Received: 14 June 2019 * Accepted:
17 February 2020 * Published: 09 March 2020 * DOI: https://doi.org/10.1038/s41598-020-61147-3 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content:
Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative
Trending News
Spatio-temporal variation of male sterile frequencies in two natural populations of beta maritimaABSTRACT The spatio-temporal variation of sex phenotype frequencies is investigated within two gynodioecious populations...
Video. Danish pm gets the giggles during statement on circus elephantsDenmark's Prime Minister Mette Frederiksen roared with laughter when giving a speech during the parliament session ...
The power at DWP - Los Angeles Times‘Nothing’s going on here,” Board of Water and Power Commissioners President Lee Kanon Alpert insisted at Tuesday’s meeti...
2 high street, purfleet, essex rm19 1qb: cam/00kg/hmf/2022/00182 HIGH STREET, PURFLEET, ESSEX RM19 1QB: CAM/00KG/HMF/2022/0018 Eastern Region, Judge K Saward and Mr J Francis QPM on 1...
Built In Brooklyn: Atavist Is Both Publisher And Platform For Online StorytellingWe returned to Brooklyn’s Dumbo neighborhood for the latest episode of Built in Brooklyn. This time we visited the offic...
Latests News
Phosphorus (p) use efficiency in rice is linked to tissue-specific biomass and p allocation patternsABSTRACT Phosphorus (P) is a non-renewable resource which may be depleted within next few decades; hence high P use effi...
Trump and the tories: a warning | thearticleAs New Hampshire appears to be yet another step along the road to the Republican nomination for Donald Trump, it’s worth...
One last hurdle remains between garrett wilson and jets blissGarrett Wilson’s megawatt smile was back in place on Wednesday when he did a 14-minute interview session with reporters....
The page you were looking for doesn't exist.You may have mistyped the address or the page may have moved.By proceeding, you agree to our Terms & Conditions and our ...
how to sign up online for parts a and b of medicareYou’re eligible for Medicare at 65, but enrollment isn’t always automatic. If you’ve received Social Security retirement...