The mammalian target of rapamycin and dna methyltransferase 1 axis mediates vascular endothelial dysfunction in response to disturbed flow
The mammalian target of rapamycin and dna methyltransferase 1 axis mediates vascular endothelial dysfunction in response to disturbed flow"
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT The earliest atherosclerotic lesions preferentially develop in arterial regions experienced disturbed blood flow, which induces endothelial expression of pro-atherogenic genes and
the subsequent endothelial dysfunction. Our previous study has demonstrated an up-regulation of DNA methyltransferase 1 (DNMT1) and a global hypermethylation in vascular endothelium
subjected to disturbed flow. Here, we determined that DNMT1-specific inhibition in arterial wall ameliorates the disturbed flow-induced atherosclerosis through, at least in part, targeting
cell cycle regulator cyclin A and connective tissue growth factor (CTGF). We identified the signaling pathways mediating the flow-induction of DNMT1. Inhibition of the mammalian target of
rapamycin (mTOR) suppressed the DNMT1 up-regulation both _in vitro_ and _in vivo_. Together, our results demonstrate that disturbed flow influences endothelial function and induces
atherosclerosis in an mTOR/DNMT1-dependent manner. The conclusions obtained from this study might facilitate further evaluation of the epigenetic regulation of endothelial function during
the pathological development of atherosclerosis and offer novel prevention and therapeutic targets of this disease. SIMILAR CONTENT BEING VIEWED BY OTHERS 2′–5′ OLIGOADENYLATE
SYNTHETASE‑LIKE 1 (OASL1) PROTECTS AGAINST ATHEROSCLEROSIS BY MAINTAINING ENDOTHELIAL NITRIC OXIDE SYNTHASE MRNA STABILITY Article Open access 04 November 2022 RECIPROCAL REGULATION OF
ENDOTHELIAL–MESENCHYMAL TRANSITION BY MAPK7 AND EZH2 IN INTIMAL HYPERPLASIA AND CORONARY ARTERY DISEASE Article Open access 07 September 2021 FLOW-INDUCED REPROGRAMMING OF ENDOTHELIAL CELLS
IN ATHEROSCLEROSIS Article 24 May 2023 INTRODUCTION The earliest atherosclerotic lesions typically originate in a nonrandom pattern, developing preferentially at arterial branches and
curvatures, where unstable disturbed blood flow is prevalent1. Shear stress resulting from disturbed flow is low and oscillates back and forth. Disturbed flow induces a dysfunctional
endothelial cell (EC) phenotype that initiates and perpetuates pro-atherogenesis1,2. In contrast, the straight parts of the arteries are protected by pulsatile unidirectional laminar flow,
and are therefore relatively atheroresistant. Vascular ECs in these regions are exposed to laminar and pulsatile shear stress that induces expressions of atheroprotective genes in the
cells1,2. Emerging evidence from our laboratory and others has revealed that epigenetic mechanisms, including DNA methylation, histone modifications, and noncoding RNA-based
post-transcriptional regulations play critical roles in the flow-regulation of endothelial gene expression and phenotype3,4,5,6,7,8,9. Of these, DNA methyltransferase (DNMT) -1 and -3A were
reported to be up-regulated by disturbed flow to promote a proatherogenic phenotype in vascular ECs3,4,8. However, the signaling pathways through which disturbed flow up-regulates DNMTs are
unclear. The mechanism by which DNMTs cause changes in EC gene expression and function is still lacking. Mapping of the mechanotransduction networks that can be activated by shear stress in
vascular ECs has indicated a series of signaling pathways as well as their critical nodes, including integrins10,11, the mitogen-activated protein kinases (MAPKs)12,13, and the mammalian
target of rapamycin (mTOR)14,15. By examining ECs in a parallel-plate flow channels exposing to pulsatile shear (PS) that mimics atheroprotective flow or oscillatory shear (OS) that mimics
atherogenic flow, our previous studies demonstrated that OS induces sustained phosphorylation of mTOR and its effector ribosomal protein S6 kinase (p70S6K)14. Integrins, whose activation
induces their associations with focal adhesion kinase (FAK) and Shc, an adaptor protein containing a C-terminal Src homology domain-2 (SH2) domain, and activation of downstream MAPK ERK1/2,
have been indicated as mechanosensors in the flow-activation of phosphatidylinositol 3-kinase (PI3K)/mTOR/p70S6K14,16,17. There is evidence that the PI3K/mTOR/p70S6K pathway is required for
EC proliferation, survival, and migration18,19. Interestingly, treatment of ECs with 5-Aza-2′-deoxycytidine (5-Aza), an inhibitor of DNMTs, limited cell proliferation and migration20,21. On
the basis of these studies, it is speculated that mTOR might mediate the OS-induction of DNMT1, which would lead to downstream changes in gene expression to result in EC dysfunction.
Although Shc and FAK have been shown to be critical for the integrin-mediated signaling, whether integrins/Shc/FAK modulate the activation of the putative mTOR-DNMT1 axis in response to OS
remains to be determined. The atherosusceptible endothelium is characterized by elevated expression of proatherogenic genes as well as consequential aberrant proliferation, migration, and
pro-inflammatory responses1. The proatherogenic genes that can be induced by application of disturbed flow or OS to ECs include cyclin A (CCNA2)22,23 and connective tissue growth factor
(CTGF)10,24, those have been found to be highly expressed in atherosclerotic plaques25,26,27. Cyclin A promotes both G1/S and G2/M transitions of the cell cycle in somatic cells. CTGF has
been associated with proliferation and inflammation in ECs and it enhances the chemosensory adhesion and migration of monocytes into atherosclerotic lesions26,27. Promoter DNA methylation
contributes to the regulatory mechanisms of cyclin A and CTGF by various stimuli such as homocysteine21 or high glucose28. In particular, a positive correlation of cyclin A promoter
methylation with cyclin A expression is found21, whereas CTGF promoter methylation is negatively correlated with the CTGF protein level28,29. In this study we have examined the role of
DNMT1-dependent promoter methylation in the flow-regulation of EC cyclin A and CTGF expression as well as the functional consequences. We also discovered the mechanotransductive mechanisms
of OS-specific up-regulation of DNMT1. RESULTS SILENCING OF DNMT1 AMELIORATES THE OS-INDUCED ENDOTHELIAL DYSFUNCTION Our previous study demonstrated that atheroprone OS induces DNA
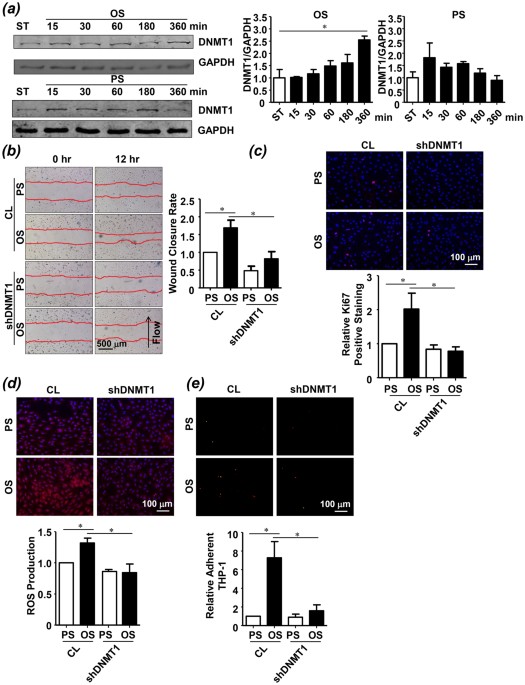
hypermethylation and DNMT1 expression in comparison to atheroprotective PS after 24 hours of shearing8. To investigate the temporal regulation of endothelial DNMT1 expression by different
patterns of shear stress, OS at 0.5 ± 4 dyn/cm2 or PS at 12 ± 4 dyn/cm2 was applied over a time course of 6 hours to human umbilical vein ECs (HUVECs). Expression of DNMT1 was induced
gradually by OS and remained elevated after 6 hours of shearing, compared with the control cells under static condition (Fig. 1a). This induction lasted over 24 hours of OS application (Fig.
S1). In contrast, a rapid induction of DNMT1 expression was observed in ECs exposed to PS; however, the induction was transient and returned to the basal level after 6 hours of shearing
(Figs 1a and S1). The followed functional experiments were therefore conducted after 6 hours of shearing unless otherwise specified. Because treatment of static ECs with non-specific DNMT
inhibitor 5-Aza suppresses migration and proliferation of the cells, we explored the mediatory role of DNMT1 in OS-induced endothelial migration and proliferation by silencing DNMT1 with
recombinant adenovirus expressing shRNA targeting human DNMT1 (ad-_h_shDNMT1). As evidenced by scratched wound healing assay and immunofluorescent staining of proliferative marker Ki67,
exposure of ECs to OS accelerated cell migration and promoted aberrant cell proliferation; these effects were abolished by DNMT1 silencing (Fig. 1b,c). These observations suggest a
pro-migrative and pro-proliferative role of DNMT1 in mediating the OS-induced endothelial dysfunction. Production of reactive oxygen species (ROS) and adhesion of circulating monocyte to
endothelium have been associated with endothelial inflammation. We found an increased ROS production and an enhanced THP-1 monocyte adhesion in ECs exposed to OS (Fig. 1d,e). Silencing of
DNMT1 suppressed the OS-induced production of ROS and decreased the relative amount of THP-1 monocytes adherent to the endothelial monolayer (Fig. 1d,e). Together, these results indicated
that DNMT1 mediates the functional modulation of OS on ECs _in vitro_. DNMT1 EXPRESSION AND GLOBAL DNA METHYLATION ARE ELEVATED IN THE ATHEROSCLEROTIC ENDOTHELIUM To gain insight into the
_in vivo_ relevance and functional implications of DNMT1 in atherosclerosis, we examined DNMT1 expression and global methylation of its substrate, DNA cytosine-5, in atherosclerotic plaques.
Apolipoprotein E-deficient (ApoE−/−) mice fed a high-fat diet (HFD) for three months and the control C57BL/6 wild-type (WT) mice were euthanized and DNMT1 and methylated 5-cytosine (5-meC)
were examined in the aortic roots. CD31 was co-stained to indicate the EC layer. The control aorta from WT mice showed weak signals of DNMT1 and 5-meC in the arterial walls (Fig. 2a,b, upper
panels; Fig. S2a,b). In contrast, a pronounced staining of DNMT1 and 5-meC was observed both in the EC layers and in the plaques (Fig. 2a,b, lower panels; Fig. S2a,b). These findings reveal
a high prevalence of DNMT1 expression and DNA hypermethylation in the atherosclerotic lesions, rather than in the undiseased regions. To explore the involvement of DNMT1 in disturbed
flow-induced endothelial dysregulation as well as atherogenesis, we studied DNMT1 expression and DNA methylation in endothelium in a mouse carotid partial ligation model, in which 3 of the 4
caudal branches of the left common carotid artery are surgically ligated (ligated), while the contralateral right common carotid artery remains untouched and serves as control (unligated).
Flow disturbance was therefore generated in the left common carotid arteries. Partial ligation in ApoE−/− mice fed a HFD resulted in atheroma formation in one month (Fig. 2c,d; Fig. S2c,d).
Consistent with observations from the three-month-HFD model, the flow-disturbance-accelerated atherosclerosis model showed up-regulated DNMT1 expression and DNA methylation in the EC layers
and in the lesion regions, in comparison with those in the normal regions (Fig. 2c,d; Fig. S2c,d). Altogether, the strong correlation between DNMT1 expression and atherosclerosis suggests a
positive contribution of DNMT1 to atherogenesis. INHIBITION OF DNMT1 AMELIORATES ENDOTHELIAL DYSFUNCTION AND ATHEROSCLEROSIS We next assessed the functional consequence of DNMT1-specific
silencing in the partial ligation model. For knockdown of vascular DNMT1, the mice received a locally intraluminal incubation of recombinant adenovirus expressing GFP and shRNA targeting
murine DNMT1 (ad-_m_shDNMT1) (Fig. 3a). Intraluminal incubation of ad-_m_shDNMT1 but not the control virus expressing GFP decreased the arterial expression of DNMT1, as evidenced by
immunofluorescent staining performed 7 days after the operation (Fig. 3b, upper panels). Staining of CD31 indicated an intact or regenerated endothelium upon virus incubation (Fig. 3b, lower
panels). An increased expression of DNMT1 in the endothelium of partially ligated arteries compared with that in the unligated was observed (Fig. 3b, upper panels). Immunofluorescence also
indicated prominent expressions of proliferating cell nuclear antigen (PCNA), vascular cell adhesion molecule 1 (VCAM1), and intercellular cell adhesion molecule 1 (ICAM1), in the vessel
walls of the ligated carotid arteries with control virus, in comparison with those in the right carotid arteries without operation and with those subjected to ad-_m_shDNMT1 incubation (Fig.
3c). Nuclear expression of PCNA reveals cell proliferation while the endothelial adhesion molecules VCAM1 and ICAM1 facilitate the accumulation of circulating monocytes on the endothelium.
These results suggest that DNMT1 promotes endothelial proliferation and inflammation _in vivo_. Moreover, we investigated the effects of DNMT1 silencing by ad-_m_shDNMT1 on atheroma
formation. The ligated arteries with intraluminal incubation of control virus rapidly developed atherosclerosis within 4 weeks of operation, and this was markedly attenuated in the arteries
with incubation of ad-_m_shDNMT1, as quantified by intima-to-media ratio and plaque area (Fig. 3d,e). Taken together, these findings provide strong evidence that DNMT1 silencing prevents
atherosclerosis by reducing endothelial proliferation and pro-inflammation in the arterial wall. MTOR/P70S6K MEDIATES THE OS-INDUCTION OF DNMT1 We investigated the molecular mechanisms
underlying the OS-induction of DNMT1. ECs were pre-incubated with mTORC1 inhibitor, rapamycin, at 50 nM for 12 hours after seeding onto the collagen-coated glass slides, and then exposed to
either PS or OS for 6 hours. Rapamycin blocked the OS-induced phosphorylation of mTOR’s effector, p70S6K, as well as the OS-induction of DNMT1 expression, as compared with the cells
pretreated with control reagent, DMSO (Fig. 4a, upper panels). Pre-treating ECs with wortmannin (100 nM, 12 hours), which is known to target PI3K/mTOR cascade upstream of p70S6K, also
suppressed the OS-induction of DNMT1 (Fig. 4a, upper panels). Coinciding with these results from experiments of pharmacological inhibition, knockdown of mTOR expression by siRNA silencing
blocked the DNMT1 up-regulation (Fig. 4a, lower left panels). Inhibition of DNMT1 by ad-_h_shDNMT1 did not affect the OS-activation of p70S6K, suggesting no feedback control of mTOR/p70S6K
signaling by DNMT1 (Fig. 4a, lower right panels). In our previous study we found that application of OS to ECs induced ERK1/2 MAPK phosphorylation, which directed the activation of mTOR14.
Therefore we tested the involvement of ERK1/2 activation in the shear stress-regulation of DNMT1. We observed that the OS-induced p70S6K phosphorylation and DNMT1 expression were inhibited
by pretreating ECs with PD98059 (50 nM), an inhibitor of MAPK kinase (MEK) that is upstream of ERK, or by transfecting the cells with siRNAs targeting ERK1 and ERK2 (Fig. 4b). To explore the
mechanosensors that perceive shear stress and translate this stimulus into biochemical signals to induce DNMT1 expression, ECs were transfected with siRNA targeting integrin β3 (siITGB3, 40
nM) or the control scrambled siRNA (siCL), and then exposed to either PS or OS. Knockdown of integrin β3 inhibited the OS-induced phosphorylation of ERK1/2 and p70S6K, and expression of
DNMT1 (Fig. 4c, upper panels). Transfecting ECs with a dominant-negative mutant of Shc (Shc-mut) but not with the wild typo control (Shc-wt) also resulted in inhibitions in OS-induced ERK1/2
and p70S6K phosphorylation as well as DNMT1 expression (Fig. 4c, lower panels), confirming the involvement of integrin and its adaptor protein, Shc, in mediating the shear-initiated
ERK/mTOR/DNMT1 signaling. Integrin-regulated protein focal adhesion kinase (FAK) has been associated with shear-activation of MAPK. We verified the contribution of FAK in directing the
OS-induced mTOR activation and DNMT1 expression by pre-incubating ECs with a FAK inhibitor or the control reagent, DMSO, and then exposed the cells to OS or PS. FAK inhibition suppressed the
OS-induction of p70S6K phosphorylation and DNMT1 expression (Fig. 4d, upper panels). Transfection of cells with siRNA targeting FAK also resulted in the suppression of OS-induced DNMT1
expression (Fig. 4d, lower panels). In summary, these _in vitro_ studies demonstrate the importance of integrin-MAPK-mTOR cascade in mediating the shear stress-induction of DNMT1 expression.
Evidence from _in vivo_ experiments confirmed this signaling mechanism by showing that the endothelial expression of DNMT1 in the ligated left carotid arteries of WT mice was inhibited by
repetitive intraperitoneal injection of rapamycin (Fig. 4e). The _in vitro_ and _in vivo_ studies strongly support a signaling model as summarized in the schematic diagram (Fig. 4f). DNMT1
TARGETS CYCLIN A AND CTGF We then searched for unidentified flow-sensitive genes whose expressions are differentially regulated by OS versus PS in a DNMT1-dependent manner. Candidates for
screening included cyclin A, CTGF, endothelial nitric oxide synthase (eNOS), Kruppel-like factor 2 (KLF2), KLF4, and others. Of these, cyclin A and CTGF showed robust up-regulation upon OS
exposure, and the up-regulation could be completely prevented by infecting the cells with ad-_h_shDNMT1 or by pretreating the cells with 5-Aza (Fig. 5a). The OS induced cell proliferation
and THP-1 adhesion could be suppressed by siRNA-mediated silencing of cyclin A and CTGF, respectively (Fig. S3). Mechanistically, OS increased the binding of DNMT1 to the promoter regions of
cyclin A, whereas it reduced DNMT1 binding to the promoter regions of CTGF (Fig. 5b), as evidenced by chromatin immunoprecipitation assay. Because DNMT1 catalyzes DNA primarily at CpG
islands within the promoter regions, we analyzed the DNA sequence at the 5′-flanking regions of human cyclin A and CTGF genes using a CpG island search program, and detected CpG-rich regions
(GC content > 50%) that span positions −291 to −100, and −153 to +46, around the promoters of the cyclin A and CTGF genes, respectively (Fig. 5c). We used methylation-specific PCR (MSP)
analysis to study promoter methylation of cyclin A and CTGF genes. Application of OS to ECs resulted in hypermethylation of cyclin A promoter and hypomethylation of CTGF promoter, in
comparison with application of PS to the cells (Fig. 5d). These observations are in line with the shear stress-regulated DNMT1 binding to the cyclin A and CTGF promoters. However, the
decrease of DNMT1 in DNA binding ability to CTGF promoter is apparently conflicting with the increase in DNMT1 expression upon OS stimulation. Therefore we next employed gain- and
loss-of-function approaches to study the correlation between expressions of DNMT1 and CTGF in static condition. Expression of cyclin A was also measured in parallel experiments. Western blot
analysis revealed that cyclin A was down-regulated by DNMT1 inhibition and was up-regulated by DNMT1 overexpression; and that CTGF expression was negatively correlated to the DNMT1
expression (Fig. 5e). Given that OS induced CTGF expression and this induction was dependent on DNMT1 (Fig. 5a), these results implied that overexpressing DNMT1 in static condition solely
could not mimic OS. Targeting of cyclin A and CTGF by DNMT1 was verified _in vivo_ by immunofluorescent staining of cyclin A and CTGF in the ligated mouse carotid arteries with or without
intraluminal silencing of DNMT1. For both the one-week and four-week models, the ligated arterials had increased levels of cyclin A and CTGF in the vessel wall, in particular the
endothelium, in comparison with the unligated (Fig. 5f,g). The increase was suppressed in the arteries with DNMT1 inhibition (Fig. 5f,g). DISCUSSION DNMTs have been shown to play important
roles in cardiovascular biology, in particular, the regulation of endothelial lineage and homeostasis30,31,32. Recent studies demonstrated that DNMT1is aberrantly expressed in endothelial
cells in response to various stimuli, including atheroprone hemodynamic shear stress3,8. Our previous report and present findings also support the pro-atherogenic role of DNMT1 in vascular
ECs subjected to low and oscillating shear stress. This study specifies the function of endothelial DNMT1 to be pro-proliferative, pro-migratory, and pro-inflammatory (Fig. 1b–e). The
conclusions we obtained _in vitro_ were further supported by the _in vivo_ results that in diseased arterial walls, DNMT1 was dysregulated and highly expressed (Fig. 2a,c). Accordingly, DNAs
from the atherosclerotic plaques were hypermethylated in comparison with those from the plaque-free regions (Fig. 2b,d). Although there is seemingly contradictory evidence in the literature
revealing that demethylation of normally hypermethylated CpG islands occurs in human atherosclerotic arteries33,34, recent study using genome-wide DNA methylation profiling at
single-nucleotide resolution indicated that DNA hypermethylation is as an overlapping feature of human atherosclerosis and an ultimate outcome of atherosclerosis-related stimuli35.
Explaining this controversy is difficult, because none of these studies considered dissecting the contribution of ECs to hypermethylation or hypomethylation. Our results are in agreement
with the notion of “global DNA hypermethylation” in atherosclerotic plaques. In particular, we attribute the gross hypermethylation, at least in part, to the endothelial up-regulation of
DNMT1 and endothelial hypermethylation. Moreover, this presented study may facilitate the establishment of a causal relationship between expression of endothelial DNMT1 and the
pro-atherogenic endothelial dysfunction and offer opportunities for therapeutic intervention of atherosclerosis. The latter is further supported by the compelling evidence from intraluminal
DNMT1 inhibition in the partial ligation model, that silencing of vascular DNMT1 suppressed the disturbed flow-induced expressions of proliferating marker PCNA and inflammatory molecules
VCAM1 and ICAM1, and attenuated the flow-disturbance-augmented atherosclerosis (Fig. 3c–e). The mechanotransduction processes depend on the mode of shearing, thus the mechano-chemical
signalings have been studied by comparing the effects of no-shearing, shearing with a significant forward direction (whether laminar shear without oscillation36 or PS in which an oscillatory
component is superimposed on a steady shearing), and shearing without a significant forward direction or with little net forward component, e.g., OS. In this study we directly compared the
effects of atheroprotective PS and atheroprone OS loading on ECs. By comparison, many other studies reported the effects of shear stress with certain flow pattern when referenced to a static
condition. However, the no-shearing condition does not exist _in vivo_ because of the existence and variation of flow velocity and pressure in the vessels. We also conducted time-course
experiments to investigate the flow-regulation of DNMT1 expression within 6 hours of shearing. DNMT1 was transiently induced by PS, by contrast, was gradually and sustainedly induced by OS
(Fig. 1a). The transient changes of gene expression upon PS exposure may be caused by shifting instantaneously from static to shear condition. At 6 hours of shearing, the majority of cells
have recovered from the adaptive responses and the OS-specific induction of DNMT1 becomes prominent. As summarized in Fig. 4f, we have demonstrated a novel mechanism by which OS specifically
up-regulates DNMT1. Our data suggests that OS-induction DNMT1 is dependent on integrin β3 (Fig. 4c), whose activation is then transmitted into cells through the phosphorylation of
integrin-associated proteins such as Shc and FAK (Fig. 4c,d). Shc and FAK signaling has been shown to be critical for the shear stress-activation of MAPKs14. Our results are consistent with
the previous findings and indicate that ERK1/2 plays an important role in mediating the activation of mTOR and the subsequence DNMT1 up-regulation (Fig. 4b). The role of mTOR signaling in
modulating DNMT1 expression in cells is controversial. The level of DNMT1 was higher after treatment of the hepatocellular carcinoma cell lines with an mTOR inhibitor, Torin-2, whereas the
increase in DNMT1expression by TGF-β treatment was abolished in fibroblasts by FAK, PI3K, and mTOR inhibitors, including rapamycin37,38. It is possible that the effects of activating mTOR on
DNMT1 expression are diverse with respect to the cell types and stimuli. Our findings showed that treatment of ECs with mTOR inhibitor rapamycin or PI3K inhibitor wortmannin blocked the
OS-induction of DNMT1 (Fig. 4a), indicating that PI3K/mTOR activations are necessary for DNMT1 up-regulation in response to OS. The mediators between mTOR/p70S6K and DNMT1 warrant further
characterizations. Cyclins associate with cyclin-dependent protein kinases (CDKs) to regulate their activity and the progression of the cell cycle. Cyclin A is highly expressed in
atherosclerosis25. We have previously demonstrated that OS promotes aberrant EC proliferation and cell cycle progression through up-regulation of cyclin A expression14. Research from others
indicated that the EC expression of cyclin A could be inhibited by homocysteine through down-regulating DNMT1 and the subsequent hypomethylation of cyclin A promoter21. Our current study
showed that OS increases DNMT1 to methylate the cyclin A promoter and therefore increases cyclin A expression (Fig. 5), confirming and extending the observations from ours and others. Our
findings also support the notion that DNA methylation on promoters can activate gene transcription, probably through the cooperation with other epigenetic machineries such as histone
acetylates/deacetylates, histone methyltransferases/demethylases, and chromatin remodeling complexes. In previous reports, pro-atherogenic CTGF has been suggested to be increased by
disturbed flow but not by steady laminar flow in ECs10,24. Here we identified an epigenetic modulator, DNMT1, which mediates the disturbed flow-induction of CTGF. Similar to the case of
cyclin A that is regulated by DNMT1, CTGF expression is positively correlated to DNMT1 expression under shearing conditions, _in vitro_ and _in vivo_ (Fig. 5a,f and g). However, the positive
correlation does not exist in static condition (Fig. 5e). Besides, OS reduced the binding of DNMT1 to CTGF promoter as well as the methylation of CTGF promoter (Fig. 5b,d). One possible
explanation for the contradictory observations between the increased expression and decreased site-specific binding of DNMT1 in response to OS may be that some co-factors are required for
recruiting or competing with DNMT1 to bind the CTGF promoter. It is likely that OS regulates the co-factors to dominate the DNMT1 binding. Further investigation will be required to
characterize the detailed mechanism by which OS induces a site-specific binding and distribution of DNMT1 along the genome. METHODS CELL CULTURE AND SHEAR EXPERIMENTS Human umbilical vein
endothelial cells (HUVECs) within passages 5–8 were maintained in Medium 199 (Gibco) supplemented with 10% fetal bovine serum (FBS) (Gibco). Bovine artery endothelial cells (BAECs) within
passages 5–18 were maintained in Medium DMEM (Gibco) supplemented with 10% fetal bovine serum (FBS) (Gemini). A parallel-plate flow apparatus was used to impose fluid shear stress to the
cells seeded on collagen I (50 μg/mL)-coated glass slides, as described previously14. The shear stress (τ) generated on the endothelial monolayer was estimated as 6Qμ/wh2, where Q is flow
rate and μ is perfusate viscosity. ECs were exposed to either pulsatile shear (PS, 12 ± 4 dynes/cm2) or oscillatory shear (OS, 0.5 ± 4 dynes/cm2). FLUORESCENT IMMUNOCYTOCHEMISTRY ECs on
glass slides were fixed in 4% paraformaldehyde for 15 minutes, permeabilized with cold PBS containing 0.4% Triton X-100 for 10 minutes, and incubated with blocking buffer (3% bovine serum
albumin in PBS) for 1 hour before incubation overnight with primary antibodies against 5-methylcytosine (5-meC) (Eurogentec), CD31(Santa Cruz Biotech) or DNMT1 (Santa Cruz Biotech). For
5-meC staining, the permeabilized cells were denatured with 2 N HCl and neutralized with 100 mM Tris-HCl (pH 8.5) before blocking. The cells were washed, incubated with secondary antibodies,
and mounted in fluorescent mounting medium with DAPI. The slides/slips were then visualized by epi–fluorescence microscopy. WESTERN BLOT ECs were lysed in the RIPA lysis buffer: 25 mM
HEPES, pH 7.4, 1% Triton X-100, 1% deoxycholate, 0.1% SDS, 125 mM NaCl, 5 mM EDTA, 50 mM NaF, 1 mM PMSF. Equal amounts of protein were separated on SDS-PAGE, transferred to nitrocellulose
membranes, blocked with 5% skim milk TBST, and incubated with the primary antibodies against DNMT1 (Santa Cruz Biotech), p-ERK (Bioworld), ERK (Bioworld), p-p70S6K (Santa Cruz Biotech),
p70S6K (Proteintech), p-FAK (Santa Cruz Biotech), FAK (Proteintech) or GAPDH (Santa Cruz Biotech). TISSUE PREPARATION, HEMATOXYLIN AND OIL RED O STAINING, AND IMMUNOFLUORESCENCE Frozen
blocks containing the hearts, aortic arches, and carotid arteries were prepared in TissueTek and stored at −80 °C. Sections from the blocks were stained with Oil red O by fixing of the
frozen tissues with 10% formalin for 10 minutes, rinsing twice with distilled water for 5 minutes, orbital shaking in 60% propanediol for 10 minutes, and incubation in 0.2% Oil red O
solution for 5 minutes at room temperature. For hematoxylin staining, the previous method was followed by 2 rinses with distilled water, a hematoxylin dip for 20 seconds, 2 rinses with
distilled water and then mounting. Immunofluorescence was performed on sections from the frozen blocks using the following antibodies at a 1:150 dilution: DNMT1 (Santa Cruz Biotech), PCNA
(Santa Cruz Biotech), VCAM1 (Santa Cruz Biotech), ICAM1 (Santa Cruz Biotech), cyclin A (Santa Cruz Biotech),CTGF (Santa Cruz Biotech), CD31 (Santa Cruz Biotech). MONOCYTE ADHESION ASSAY ECs
were pre-incubated with adenovirus and were subjected to either PS or OS for 6 hours. Human peripheral blood mononuclear leukocytes (THP-1 cells) were cultured in RPMI 1640 medium
(Invitrogen) and were labeled in serum-free medium with CM-Dil (1 g/ml) at 37 °C for 30 minutes, followed by at 4 °C for 10 minutes. After shearing, ECs were incubated with the labeled THP-1
cells at a concentration of 5 × 105 THP-1 cells per milliliter (in a total of 4 ml) for 30 minutes at 37 °C. Non-adherent monocytes were washed away with 1640, and the adherent monocytes
were fixed with 4% paraformaldehyde for 5 minutes. Cells were counted in 10 randomly selected microscopic fields under an inverted epi-fluorescence microscope. CELL PROLIFERATION MEASURED BY
KI67 STAINING Standard fluorescent immunocytochemistry was employed to assay cell proliferation, as indicated by a positive staining of proliferative marker, Ki67. The numbers of stained
cell nuclei were scored in 10 randomly selected microscopic fields and the ratios of stained to total cells (DAPI-positive cells) expressed as numerical value relative to the CL/PS were
defined as Ki67 positive staining. METHYLATION SPECIFIC-POLYMERASE CHAIN REACTION (MSP) Genomic DNA was extracted for methylation analysis from cells after exposure to PS or OS for 6 hours.
One gram of genomic DNA was modified with sodium bisulfite using the EpiMark Bisulfite kit (BioLabs) according to the manufacturer’s instructions. Polymerase chain reaction (PCR)
amplifications were carried out in a total volume of 25 μL by using 2× EasyTaq PCR SuperMix (TRANSGEN BIOTECH). The primer sets are listed in Table S1. MSP reactions were subjected to
initial incubation at 95 °C for 5 minutes, followed by 40 cycles of 95 °C for 30 seconds, and annealing at the appropriate temperature for 30 seconds and 72 °C for 30 seconds. Final
extension was done by incubation at 72 °C for 5 minutes. MSP products were separated on 2% agarose gels and visualized after Gel-Red (Beyotime) staining. CAROTID PARTIAL LIGATION AND VESSEL
HARVESTING All animal studies were performed in accordance with the approved protocol of the Animal Care and Use Committee of Peking University and were performed in accordance with the
“Guide for the care and use of laboratory animals” published by the US National Institutes of Health (publication No. 85–23, revised 1996). The surgical procedures are modified from the
previous reports3,8. Briefly, the left carotid bifurcation of mice was exposed following a neck incision. Three branches (external carotid, internal carotid, and occipital) of the left
carotid artery were ligated with a 6-0 silk suture, and the superior thyroid artery was left intact. The right carotid arteries were served as control. At one week after operation, the
animals were sacrificed and the vessels were perfused with a fixative (4% paraformaldehyde in PBS) under pressure (100 mmHg). The arteries were dissected out and further fixed by immersion
in the fixative for 16 hours before being embedded in Tissue-Tek OCT compound and stored in −80 °C. _IN VIVO_ DELIVERY OF VIRUS ApoE−/− and C57BL/6 wildtype mice (8–12 weeks old, 18–25 g)
were anesthetized and were then subjected to partial ligation of carotid artery. After the surgery, some mice were injected intraperitoneally twice per week with rapamycin (1 mg/kg/day) or
with control reagent. Some mice received a locally intraluminal incubation of ad-_m_shDNMT1 (Gene Pharma) or control virus. Briefly, left common carotid artery was exposed by blunt
dissection. The internal carotid and occipital artery were ligated with 6–0 silk suture. Heart proximal end of the left common carotid artery was clamped temporarily. Adenovirus was injected
into the common carotid artery through external carotid artery with a catheter and was retained intraluminally in the artery for 30 minutes. After that the external carotid artery was
ligated while the superior thyroid artery was left intact. The clamp was removed. ApoE−/− mice were fed with a Western diet (D12108C high fat rodent diet with 1.25% cholesterol) immediately
after surgery. One or four weeks after ligation, the mice were sacrificed and fixed for 5 minutes by perfusion through left cardiac ventricle with 4% paraformaldehyde in PBS under
physiological pressure. The ligated carotid arteries were harvested and subjected to histology and immunostaining analyses of the vessels. MIGRATION ASSAY EC migration was evaluated with
scratch wound healing assay. In brief, the cells were subjected to either PS or OS for 6 hours. Then the monolayer was scratched using a pipette tip, and the cells were then allowed to
migrate for 12 hours. Wound closure rate of the scratch was measured to quantitatively evaluate cell migration. (Wound closure rate = (A0 − A t) /A0 × 100%, A0: mean width of scratches at 0
hour, At: mean width of scratches at 12 hours). ROS PRODUCTION ASSAY ROS production was detected with DHE kit (KeyGEN BioTECH) according to the manufacturer’s instructions. In brief, ECs
were subjected to either PS or OS for 6 hours, and then the cells were incubated in DHE (25 μM in M199 basal medium) for 30 minutes at room temperature. The cells were then visualized by
epi–fluorescence microscopy. CHROMATIN IMMUNOPRECIPITATION ECs were subjected to either PS or OS for 6 hours. After shearing, the cells were washed once with PBS (room temperature), fixed
with 1% formaldehyde in PBS for 10 minutes, rinsed twice with ice-cold PBS, and scraped into 1 ml of ice-cold PBS. The pellet was resuspended with 300 μl of lysis buffer (1% SDS, 5 mM EDTA,
50 mM Tris-HCl [pH 8.1], and protease inhibitors), incubated on ice for 10 minutes, and sonicated for 3 times at 12 seconds each. A 10% aliquot was saved as an input. The lysate was 1:10
diluted in dilution buffer (1% Triton X-100, 2 mM EDTA, 150 mM NaCl, 20 mM Tris-HCl [pH 8.1], and protease inhibitors), and was incubated with antibody against DNMT1 (Santa Cruz Biotech) for
6 hours or overnight at 4 °C and then with 30 μl of protein A-G sepharose beads for another 2 hours. The sepharose beads were washed sequentially with buffer TSE I (0.1% SDS, 1% Triton
X-100, 2 mM EDTA, 20 mM Tris.HCl, pH 8.1, 150 mM NaCl), buffer TSE II (0.1% SDS, 1% Triton X-100, 2 mM EDTA, 20 mM Tris.HCl, pH 8.1, 500 mM NaCl), buffer III (0.25 M LiCl, 1% NP-40, 1%
deoxycholate, 1 mM EDTA, 10 mM Tris.HCl, pH 8.1), and TE buffer. The beads were then treated with RNase A (50 μg/mL) and proteinase K (7.5 μL of 20 mg/mL) at 37 °C for 30 minutes.
Cross-links were reversed at 65 °C overnight. DNA was extracted with DNA Pure-Spin Kit(Vigorous), and subjected for PCR amplification. The primer sets are listed in Table S1. SIRNA-MEDIATED
GENE SILENCING ECs were transfected with lipofectamine 2000 reagent according to the manufacturer’s instructions. The sequences of siRNAs are listed in Table S1. STATISTICAL ANALYSIS Data
are expressed as mean ± SEM from at least 3 independent experiments. Statistical analysis was performed by unpaired or paired t-test for 2 groups of data and by 1-way ANOVA for multiple
comparisons. Statistical significance among multiple groups was determined by post hoc analysis (Tukey honestly significant difference test). Values of P < 0.05 were considered
statistically significant. DATA AVAILABILITY STATEMENT The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable
request. REFERENCES * Chiu, J. J. & Chien, S. Effects of disturbed flow on vascular endothelium: pathophysiological basis and clinical perspectives. _Physiol Rev_ 91(1), 327–387 (2011).
Article PubMed MathSciNet Google Scholar * Zhou, J., Li, Y. S. & Chien, S. Shear stress-initiated signaling and its regulation of endothelial function. _Arterioscler Thromb Vasc
Biol_ 34(10), 2191–2198 (2014). Article CAS PubMed PubMed Central Google Scholar * Dunn, J. _et al_. Flow-dependent epigenetic DNA methylation regulates endothelial gene expression and
atherosclerosis. _J Clin Invest_ 124(7), 3187–3199 (2014). Article CAS PubMed PubMed Central Google Scholar * Jiang, Y. Z. _et al_. Hemodynamic disturbed flow induces differential DNA
methylation of endothelial Kruppel-Like Factor 4 promoter _in vitro_ and _in vivo_. _Circ Res_ 115(1), 32–43 (2014). Article CAS PubMed PubMed Central Google Scholar * Lee, D. Y. _et
al_. MicroRNA-10a is crucial for endothelial response to different flow patterns via interaction of retinoid acid receptors and histone deacetylases. _Proc Natl Acad Sci USA_ 114(8),
2072–2077 (2017). Article CAS PubMed PubMed Central Google Scholar * Zhou, J. _et al_. MicroRNA-21 targets peroxisome proliferators-activated receptor-alpha in an autoregulatory loop to
modulate flow-induced endothelial inflammation. _Proc Natl Acad Sci USA_ 108(25), 10355–10360 (2011). Article ADS CAS PubMed PubMed Central Google Scholar * Illi, B. _et al_. Shear
stress-mediated chromatin remodeling provides molecular basis for flow-dependent regulation of gene expression. _Circ Res_ 93(2), 155–161 (2003). Article CAS PubMed Google Scholar *
Zhou, J., Li, Y. S., Wang, K. C. & Chien, S. Epigenetic Mechanism in Regulation of Endothelial Function by Disturbed Flow: Induction of DNA Hypermethylation by DNMT1. _Cell Mol Bioeng_
7(2), 218–224 (2014). Article CAS PubMed PubMed Central Google Scholar * Huang, T. S. _et al_. LINC00341 exerts an anti-inflammatory effect on endothelial cells by repressing VCAM1.
_Physiol Genomics_, physiolgenomics 00132 02016 (2017). * Wang, L. _et al_. Integrin-YAP/TAZ-JNK cascade mediates atheroprotective effect of unidirectional shear flow. _Nature_ (2016). *
Sun, X. _et al_. Activation of integrin alpha5 mediated by flow requires its translocation to membrane lipid rafts in vascular endothelial cells. _Proc Natl Acad Sci USA_ 113(3), 769–774
(2016). Article ADS CAS PubMed PubMed Central Google Scholar * Shaik, S. S. _et al_. Low intensity shear stress increases endothelial ELR+ CXC chemokine production via a focal adhesion
kinase-p38{beta} MAPK-NF-{kappa}B pathway. _J Biol Chem_ 284(9), 5945–5955 (2009). Article CAS PubMed PubMed Central Google Scholar * Chiu, J. J. _et al_. Mechanisms of induction of
endothelial cell E-selectin expression by smooth muscle cells and its inhibition by shear stress. _Blood_ 110(2), 519–528 (2007). Article CAS PubMed PubMed Central Google Scholar *
Zhou, J. _et al_. BMP receptor-integrin interaction mediates responses of vascular endothelial Smad1/5 and proliferation to disturbed flow. _J Thromb Haemost_ 11(4), 741–755 (2013). Article
CAS PubMed Google Scholar * Li, B., Zhang, J., Wang, Z. & Chen, S. Ivabradine Prevents Low Shear Stress Induced Endothelial Inflammation and Oxidative Stress via mTOR/eNOS Pathway.
_PLoS One_ 11(2), e0149694 (2016). Article PubMed PubMed Central CAS Google Scholar * Lee, D. Y. _et al_. Oscillatory flow-induced proliferation of osteoblast-like cells is mediated by
alphavbeta3 and beta1 integrins through synergistic interactions of focal adhesion kinase and Shc with phosphatidylinositol 3-kinase and the Akt/mTOR/p70S6K pathway. _J Biol Chem_ 285(1),
30–42 (2010). Article CAS PubMed Google Scholar * Obi, S. _et al_. Fluid shear stress induces differentiation of circulating phenotype endothelial progenitor cells. _Am J Physiol Cell
Physiol_ 303(6), C595–606 (2012). Article CAS PubMed Google Scholar * Matter, C. M. _et al_. Effects of tacrolimus or sirolimus on proliferation of vascular smooth muscle and endothelial
cells. _J Cardiovasc Pharmacol_ 48(6), 286–292 (2006). Article CAS PubMed Google Scholar * Wang, Y., Chen, J., Tang, W., Zhang, Y. & Li, X. Rapamycin inhibits the proliferation of
endothelial cells in hemangioma by blocking the mTOR-FABP4 pathway. _Biomed Pharmacother_ 85, 272–279 (2017). Article CAS PubMed Google Scholar * Wu, J. _et al_. Kaposi’s
sarcoma-associated herpesvirus (KSHV) vIL-6 promotes cell proliferation and migration by upregulating DNMT1 via STAT3 activation. _PLoS One_ 9(3), e93478 (2014). Article ADS PubMed PubMed
Central CAS Google Scholar * Jamaluddin, M. D. _et al_. Homocysteine inhibits endothelial cell growth via DNA hypomethylation of the cyclin A gene. _Blood_ 110(10), 3648–3655 (2007).
Article CAS PubMed PubMed Central Google Scholar * Zhou, J. _et al_. Force-specific activation of Smad1/5 regulates vascular endothelial cell cycle progression in response to disturbed
flow. _Proc Natl Acad Sci USA_ 109(20), 7770–7775 (2013). Article ADS Google Scholar * Lee, D. Y. _et al_. Role of histone deacetylases in transcription factor regulation and cell cycle
modulation in endothelial cells in response to disturbed flow. _Proc Natl Acad Sci USA_ 109(6), 1967–1972 (2012). Article ADS CAS PubMed PubMed Central Google Scholar * Wang, K. C. _et
al_. Flow-dependent YAP/TAZ activities regulate endothelial phenotypes and atherosclerosis. _Proc Natl Acad Sci USA_ 113(41), 11525–11530 (2016). Article CAS PubMed PubMed Central
Google Scholar * Zettler, M. E., Merchant, M. A. & Pierce, G. N. Augmented cell cycle protein expression and kinase activity in atherosclerotic rabbit vessels. _Exp Clin Cardiol_ 15(4),
e139–144 (2010). CAS PubMed PubMed Central Google Scholar * Cicha, I. _et al_. Connective tissue growth factor is released from platelets under high shear stress and is differentially
expressed in endothelium along atherosclerotic plaques. _Clin Hemorheol Microcirc_ 35(1–2), 203–206 (2006). CAS PubMed Google Scholar * Yan, L. F. _et al_. Proliferative phenotype of
pulmonary microvascular endothelial cells plays a critical role in the overexpression of CTGF in the bleomycin-injured rat. _Exp Toxicol Pathol_ 66(1), 61–71 (2014). Article CAS PubMed
Google Scholar * Zhang, H. _et al_. High glucose-induced cytoplasmic translocation of Dnmt3a contributes to CTGF hypo-methylation in mesangial cells. _Biosci Rep_ 36 (4) (2016). * Zhang, H.
_et al_. Correlation of CTGF gene promoter methylation with CTGF expression in type 2 diabetes mellitus with or without nephropathy. _Mol Med Rep_ 9(6), 2138–2144 (2014). Article CAS
PubMed PubMed Central Google Scholar * Banerjee, S. & Bacanamwo, M. DNA methyltransferase inhibition induces mouse embryonic stem cell differentiation into endothelial cells. _Exp
Cell Res_ 316(2), 172–180 (2010). Article CAS PubMed Google Scholar * Krause, B. J. _et al_. Role of DNA methyltransferase 1 on the altered eNOS expression in human umbilical endothelium
from intrauterine growth restricted fetuses. _Epigenetics_ 8(9), 944–952 (2013). Article CAS PubMed PubMed Central Google Scholar * Dunn, J., Simmons, R., Thabet, S. & Jo, H. The
role of epigenetics in the endothelial cell shear stress response and atherosclerosis. _Int J Biochem Cell Biol_ 67, 167–176 (2015). Article CAS PubMed PubMed Central Google Scholar *
Castillo-Diaz, S. A., Garay-Sevilla, M. E., Hernandez-Gonzalez, M. A., Solis-Martinez, M. O. & Zaina, S. Extensive demethylation of normally hypermethylated CpG islands occurs in human
atherosclerotic arteries. _Int J Mol Med_ 26(5), 691–700 (2010). CAS PubMed Google Scholar * Greissel, A. _et al_. Alternation of histone and DNA methylation in human atherosclerotic
carotid plaques. _Thromb Haemost_ 114(2), 390–402 (2015). Article CAS PubMed Google Scholar * Zaina, S. _et al_. DNA methylation map of human atherosclerosis. _Circ Cardiovasc Genet_
7(5), 692–700 (2014). Article CAS PubMed Google Scholar * Wang, X. _et al_. Shear stress activation of nuclear receptor PXR in endothelial detoxification. _Proc Natl Acad Sci USA_
110(32), 13174–13179 (2013). Article ADS CAS PubMed PubMed Central Google Scholar * Wang, C. _et al_. The novel mTOR inhibitor Torin-2 induces autophagy and downregulates the
expression of UHRF1 to suppress hepatocarcinoma cell growth. _Oncol Rep_ 34(4), 1708–1716 (2015). Article CAS PubMed MathSciNet Google Scholar * Koh, H. B., Scruggs, A. M. & Huang,
S. K. Transforming Growth Factor-beta1 Increases DNA Methyltransferase 1 and 3a Expression through Distinct Post-transcriptional Mechanisms in Lung Fibroblasts. _J Biol Chem_ 291(37),
19287–19298 (2016). Article CAS PubMed PubMed Central Google Scholar Download references ACKNOWLEDGEMENTS This work was funded by the National Natural Science Foundation of the P.R.
China 81470590, 31522022, 91539116 (to J. Z.), and 81470556 (to W. P.); Beijing Natural Science Foundation 7152081 (to J. Z.). AUTHOR INFORMATION Author notes * Yun-Peng Zhang and Yi-Tao
Huang contributed equally to this work. AUTHORS AND AFFILIATIONS * Department of Physiology and Pathophysiology, School of Basic Medical Sciences, Peking University, Key Laboratory of
Molecular Cardiovascular Science, Ministry of Education, Beijing, 100191, P.R. China Yun-Peng Zhang, Yi-Tao Huang, Wei Pang, Juan-Juan Zhu, Yue-Feng Liu, Run-Ze Tang, Chuan-Rong Zhao,
Wei-Juan Yao & Jing Zhou * Department of Bioengineering and Institute of Engineering in Medicine, University of California, San Diego, La Jolla, CA, 92093, USA Tse-Shun Huang, Yi-Shuan
Li & Shu Chien Authors * Yun-Peng Zhang View author publications You can also search for this author inPubMed Google Scholar * Yi-Tao Huang View author publications You can also search
for this author inPubMed Google Scholar * Tse-Shun Huang View author publications You can also search for this author inPubMed Google Scholar * Wei Pang View author publications You can also
search for this author inPubMed Google Scholar * Juan-Juan Zhu View author publications You can also search for this author inPubMed Google Scholar * Yue-Feng Liu View author publications
You can also search for this author inPubMed Google Scholar * Run-Ze Tang View author publications You can also search for this author inPubMed Google Scholar * Chuan-Rong Zhao View author
publications You can also search for this author inPubMed Google Scholar * Wei-Juan Yao View author publications You can also search for this author inPubMed Google Scholar * Yi-Shuan Li
View author publications You can also search for this author inPubMed Google Scholar * Shu Chien View author publications You can also search for this author inPubMed Google Scholar * Jing
Zhou View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS J.Z. and Y.-P.Z. designed study. Y.-P.Z. and Y.-T.H. conducted experiments. T.-S.H.,
W.P., J.-J.Z., Y.-F.L., R.-Z.T., C.-R.Z., W.-J.Y., Y.-S.L., and S.C. discussed the data. J.Z., Y.-P.Z., and Y.-T.H. wrote the main manuscript text and prepared figures. CORRESPONDING AUTHOR
Correspondence to Jing Zhou. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare that they have no competing interests. ADDITIONAL INFORMATION PUBLISHER'S NOTE: Springer Nature
remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. ELECTRONIC SUPPLEMENTARY MATERIAL SUPPLEMENTARY INFORMATION RIGHTS AND PERMISSIONS OPEN
ACCESS This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or
format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or
other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in
the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the
copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Zhang, YP., Huang, YT.,
Huang, TS. _et al._ The Mammalian Target of Rapamycin and DNA methyltransferase 1 axis mediates vascular endothelial dysfunction in response to disturbed flow. _Sci Rep_ 7, 14996 (2017).
https://doi.org/10.1038/s41598-017-15387-5 Download citation * Received: 20 April 2017 * Accepted: 16 October 2017 * Published: 08 November 2017 * DOI:
https://doi.org/10.1038/s41598-017-15387-5 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not
currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative
Trending News
Long-term thermo-mechanical properties of the continental lithosphereABSTRACT Simple thermo-elastic models successfully explain flexure observations from oceanic lithosphere and are consist...
A model for network-based identification and pharmacological targeting of aberrant, replication-permissive transcriptional programs induced by viral iABSTRACT SARS-CoV-2 hijacks the host cell transcriptional machinery to induce a phenotypic state amenable to its replica...
How can france fight invasive species like asian hornets?ALL PROBLEM SPECIES NUMBERS HAVE GROWN DESPITE A 2017 ERADICATION PLAN AND SOME SURPRISING INVENTIONS Collective action ...
Novak djokovic opens up on roger federer and rafael nadal's us open absenceNOVAK DJOKOVIC: PUNDITS DISCUSS CORONAVIRUS STATEMENT Novak Djokovic admits it will be 'strange' not to have R...
Minerva Ornaments | NatureABSTRACT I NOTICE that a correspondent writing from America expresses his scepticism as to the figural character of cert...
Latests News
The mammalian target of rapamycin and dna methyltransferase 1 axis mediates vascular endothelial dysfunction in response to disturbed flowABSTRACT The earliest atherosclerotic lesions preferentially develop in arterial regions experienced disturbed blood flo...
How can france fight invasive species like asian hornets?ALL PROBLEM SPECIES NUMBERS HAVE GROWN DESPITE A 2017 ERADICATION PLAN AND SOME SURPRISING INVENTIONS Collective action ...
Novak djokovic opens up on roger federer and rafael nadal's us open absenceNOVAK DJOKOVIC: PUNDITS DISCUSS CORONAVIRUS STATEMENT Novak Djokovic admits it will be 'strange' not to have R...
Minerva Ornaments | NatureABSTRACT I NOTICE that a correspondent writing from America expresses his scepticism as to the figural character of cert...
Arya vidyapeeth college job recruitment 2021 for 4 assistant professor job vacancy, openingCandidates having essential qualification given above are advised to submit their applications in the DHE's prescri...