Structure of severe fever with thrombocytopenia syndrome virus l protein elucidates the mechanisms of viral transcription initiation
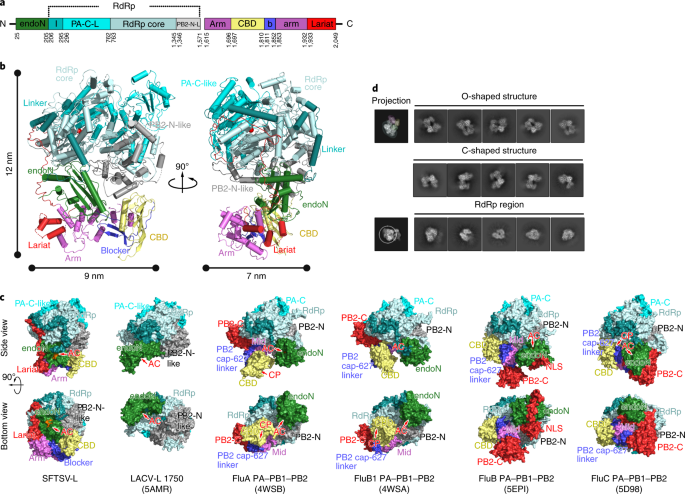
Structure of severe fever with thrombocytopenia syndrome virus l protein elucidates the mechanisms of viral transcription initiation"
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT Segmented negative-sense RNA viruses (sNSRVs) encode a single-polypeptide polymerase (L protein) or a heterotrimeric polymerase complex to cannibalize host messenger RNA cap
structures serving as primers of transcription, and catalyse RNA synthesis. Here, we report the full-length structure of the severe fever with thrombocytopaenia syndrome virus (SFTSV) L
protein, as determined by cryogenic electron microscopy at 3.4 Å, leading to an atomic model harbouring three functional parts (an endonuclease, an RNA-dependent RNA polymerase and a
cap-binding domain) and two structural domains (an arm domain with a blocker motif and a carboxy-terminal lariat domain). The SFTSV L protein has a compact architecture in which its
cap-binding pocket is surprisingly occupied by an Arg finger of the blocker motif, and the endonuclease active centre faces back towards the cap-binding pocket, suggesting that domain
rearrangements are necessary to acquire the pre-initiation state of the active site. Our results provide insight into the complete architecture of sNSRV-encoded L protein and further the
understanding of sNSRV transcription initiation. Access through your institution Buy or subscribe This is a preview of subscription content, access via your institution ACCESS OPTIONS Access
through your institution Access Nature and 54 other Nature Portfolio journals Get Nature+, our best-value online-access subscription $29.99 / 30 days cancel any time Learn more Subscribe to
this journal Receive 12 digital issues and online access to articles $119.00 per year only $9.92 per issue Learn more Buy this article * Purchase on SpringerLink * Instant access to full
article PDF Buy now Prices may be subject to local taxes which are calculated during checkout ADDITIONAL ACCESS OPTIONS: * Log in * Learn about institutional subscriptions * Read our FAQs *
Contact customer support SIMILAR CONTENT BEING VIEWED BY OTHERS STRUCTURAL INSIGHTS INTO THE RNA-DEPENDENT RNA POLYMERASE COMPLEXES FROM HIGHLY PATHOGENIC MARBURG AND EBOLA VIRUSES Article
Open access 31 March 2025 PRE-INITIATION AND ELONGATION STRUCTURES OF FULL-LENGTH LA CROSSE VIRUS POLYMERASE REVEAL FUNCTIONALLY IMPORTANT CONFORMATIONAL CHANGES Article Open access 17 July
2020 STRUCTURAL INSIGHTS INTO RNA POLYMERASES OF NEGATIVE-SENSE RNA VIRUSES Article 25 January 2021 DATA AVAILABILITY The cryo-EM density maps and structures were deposited in the Electron
Microscopy Data Bank and Protein Data Bank with the accession codes 0828 and 6L42, respectively. The data that support the findings of this study are available from the corresponding author
upon request. CHANGE HISTORY * _ 21 APRIL 2021 A Correction to this paper has been published: https://doi.org/10.1038/s41564-021-00906-y _ REFERENCES * Morin, B., Kranzusch, P. J., Rahmeh,
A. A. & Whelan, S. P. The polymerase of negative-stranded RNA viruses. _Curr. Opin. Virol._ 3, 103–110 (2013). Article CAS PubMed PubMed Central Google Scholar * Li, J., Rahmeh, A.,
Morelli, M. & Whelan, S. P. A conserved motif in region V of the large polymerase proteins of nonsegmented negative-sense RNA viruses that is essential for mRNA capping. _J. Virol._ 82,
775–784 (2008). Article CAS PubMed Google Scholar * Ogino, T. & Banerjee, A. K. Unconventional mechanism of mRNA capping by the RNA-dependent RNA polymerase of vesicular stomatitis
virus. _Mol. Cell_ 25, 85–97 (2007). Article CAS PubMed Google Scholar * Li, J., Wang, J. T. & Whelan, S. P. A unique strategy for mRNA cap methylation used by vesicular stomatitis
virus. _Proc. Natl Acad. Sci. USA_ 103, 8493–8498 (2006). Article CAS PubMed PubMed Central Google Scholar * Morin, B. et al. The N-terminal domain of the arenavirus L protein is an RNA
endonuclease essential in mRNA transcription. _PLoS Pathog._ 6, e1001038 (2010). Article PubMed PubMed Central CAS Google Scholar * Gogrefe, N., Reindl, S., Gunther, S. &
Rosenthal, M. Structure of a functional cap-binding domain in Rift Valley fever virus L protein. _PLoS Pathog._ 15, e1007829 (2019). Article CAS PubMed PubMed Central Google Scholar *
Yuan, P. et al. Crystal structure of an avian influenza polymerase PAN reveals an endonuclease active site. _Nature_ 458, 909–913 (2009). Article CAS PubMed Google Scholar * Dias, A. et
al. The cap-snatching endonuclease of influenza virus polymerase resides in the PA subunit. _Nature_ 458, 914–918 (2009). Article CAS PubMed Google Scholar * Abudurexiti, A. et al.
Taxonomy of the order Bunyavirales: update 2019. _Arch. Virol._ 164, 1949–1965 (2019). Article CAS PubMed PubMed Central Google Scholar * Guu, T. S., Zheng, W. & Tao, Y. J.
Bunyavirus: structure and replication. _Adv. Exp. Med. Biol._ 726, 245–266 (2012). Article CAS PubMed Google Scholar * Fodor, E. The RNA polymerase of influenza A virus: mechanisms of
viral transcription and replication. _Acta Virol._ 57, 113–122 (2013). Article CAS PubMed Google Scholar * Liang, B. et al. Structure of the L protein of vesicular stomatitis virus from
electron cryomicroscopy. _Cell_ 162, 314–327 (2015). Article CAS PubMed PubMed Central Google Scholar * Gilman, M. S. A. et al. Structure of the respiratory syncytial virus polymerase
complex. _Cell_ 179, 193–204 (2019). Article CAS PubMed PubMed Central Google Scholar * Pan, J. et al. Structure of the human metapneumovirus polymerase phosphoprotein complex. _Nature_
577, 275–279 (2020). Article CAS PubMed Google Scholar * Pflug, A., Guilligay, D., Reich, S. & Cusack, S. Structure of influenza A polymerase bound to the viral RNA promoter.
_Nature_ 516, 355–360 (2014). Article CAS PubMed Google Scholar * Reich, S. et al. Structural insight into cap-snatching and RNA synthesis by influenza polymerase. _Nature_ 516, 361–366
(2014). Article CAS PubMed Google Scholar * Chang, S. et al. Cryo-EM structure of influenza virus RNA polymerase complex at 4.3 A resolution. _Mol. Cell_ 57, 925–935 (2015). Article CAS
PubMed Google Scholar * Fan, H. et al. Structures of influenza A virus RNA polymerase offer insight into viral genome replication. _Nature_ 573, 287–290 (2019). Article CAS PubMed
PubMed Central Google Scholar * Hengrung, N. et al. Crystal structure of the RNA-dependent RNA polymerase from influenza C virus. _Nature_ 527, 114–117 (2015). Article CAS PubMed PubMed
Central Google Scholar * Peng, Q. et al. Structural insight into RNA synthesis by influenza D polymerase. _Nat. Microbiol._ 4, 1750–1759 (2019). Article CAS PubMed Google Scholar *
Gerlach, P., Malet, H., Cusack, S. & Reguera, J. Structural insights into bunyavirus replication and its regulation by the vRNA promoter. _Cell_ 161, 1267–1279 (2015). Article CAS
PubMed PubMed Central Google Scholar * Yu, X. J. et al. Fever with thrombocytopenia associated with a novel bunyavirus in China. _N. Engl. J. Med._ 364, 1523–1532 (2011). Article CAS
PubMed PubMed Central Google Scholar * Jiao, L. et al. Structure of severe fever with thrombocytopenia syndrome virus nucleocapsid protein in complex with suramin reveals therapeutic
potentials. _J. Virol._ 87, 6829–6839 (2013). Article CAS PubMed PubMed Central Google Scholar * Zhou, H., Sun, Y., Guo, Y. & Lou, Z. Structural perspective on the formation of
ribonucleoprotein complex in negative-sense single-stranded RNA viruses. _Trends Microbiol._ 21, 475–484 (2013). Article CAS PubMed Google Scholar * Zhou, H. et al. The nucleoprotein of
severe fever with thrombocytopenia syndrome virus processes an oligomeric ring to facilitate RNA encapsidation. _Protein Cell_ 4, 445–455 (2013). Article CAS PubMed PubMed Central Google
Scholar * Halldorsson, S. et al. Structure of a phleboviral envelope glycoprotein reveals a consolidated model of membrane fusion. _Proc. Natl Acad. Sci. USA_ 113, 7154–7159 (2016).
Article CAS PubMed PubMed Central Google Scholar * Wu, Y. et al. Structures of phlebovirus glycoprotein Gn and identification of a neutralizing antibody epitope. _Proc. Natl Acad. Sci.
USA_ 114, E7564–E7573 (2017). CAS PubMed PubMed Central Google Scholar * _2018 Annual Review of Diseases Prioritized Under the Research and Development Blueprint_ (World Health
Organization, 2018); http://www.who.int/emergencies/diseases/2018prioritization-report.pdf * Guilligay, D. et al. The structural basis for cap binding by influenza virus polymerase subunit
PB2. _Nat. Struct. Mol. Biol._ 15, 500–506 (2008). Article CAS PubMed Google Scholar * Thierry, E. et al. Influenza polymerase can adopt an alternative configuration involving a radical
repacking of PB2 domains. _Mol. Cell_ 61, 125–137 (2016). Article CAS PubMed PubMed Central Google Scholar * Knizewski, L., Kinch, L. N., Grishin, N. V., Rychlewski, L. & Ginalski,
K. Realm of PD-(D/E)XK nuclease superfamily revisited: detection of novel families with modified transitive meta profile searches. _BMC Struct. Biol._ 7, 40 (2007). Article PubMed PubMed
Central CAS Google Scholar * Reguera, J., Weber, F. & Cusack, S. Bunyaviridae RNA polymerases (L-protein) have an N-terminal, influenza-like endonuclease domain, essential for viral
cap-dependent transcription. _PLoS Pathog._ 6, e1001101 (2010). Article PubMed PubMed Central CAS Google Scholar * Wallat, G. D. et al. High-resolution structure of the N-terminal
endonuclease domain of the Lassa virus L polymerase in complex with magnesium ions. _PLoS ONE_ 9, e87577 (2014). Article PubMed PubMed Central CAS Google Scholar * Tao, Y., Farsetta, D.
L., Nibert, M. L. & Harrison, S. C. RNA synthesis in a cage—structural studies of reovirus polymerase λ3. _Cell_ 111, 733–745 (2002). Article CAS PubMed Google Scholar * Gong, P.
& Peersen, O. B. Structural basis for active site closure by the poliovirus RNA-dependent RNA polymerase. _Proc. Natl Acad. Sci. USA_ 107, 22505–22510 (2010). Article CAS PubMed
PubMed Central Google Scholar * Zheng, S. Q. et al. MotionCor2: anisotropic correction of beam-induced motion for improved cryo-electron microscopy. _Nat. Methods_ 14, 331–332 (2017).
Article CAS PubMed PubMed Central Google Scholar * Scheres, S. H. RELION: implementation of a Bayesian approach to cryo-EM structure determination. _J. Struct. Biol._ 180, 519–530
(2012). Article CAS PubMed PubMed Central Google Scholar * Rohou, A. & Grigorieff, N. CTFFIND4: fast and accurate defocus estimation from electron micrographs. _J. Struct. Biol._
192, 216–221 (2015). Article PubMed PubMed Central Google Scholar * Hu, M. et al. A particle-filter framework for robust cryo-EM 3D reconstruction. _Nat. Methods_ 15, 1083–1089 (2018).
Article CAS PubMed Google Scholar * Tan, Y. Z. et al. Addressing preferred specimen orientation in single-particle cryo-EM through tilting. _Nat. Methods_ 14, 793–796 (2017). Article
CAS PubMed PubMed Central Google Scholar * Naydenova, K. & Russo, C. J. Measuring the effects of particle orientation to improve the efficiency of electron cryomicroscopy. _Nat.
Commun._ 8, 629 (2017). Article PubMed PubMed Central CAS Google Scholar * Pettersen, E. F. et al. UCSF Chimera—a visualization system for exploratory research and analysis. _J. Comput.
Chem._ 25, 1605–1612 (2004). Article CAS PubMed Google Scholar * Jones, R. et al. Structure and function of the Toscana virus cap-snatching endonuclease. _Nucleic Acids Res._ 47,
10914–10930 (2019). Article CAS PubMed PubMed Central Google Scholar * Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. _Acta Crystallogr. D
Biol. Crystallogr._ 66, 486–501 (2010). Article CAS PubMed PubMed Central Google Scholar * Afonine, P. V. et al. Towards automated crystallographic structure refinement with
phenix.refine. _Acta Crystallogr. D Biol. Crystallogr._ 68, 352–367 (2012). Article CAS PubMed PubMed Central Google Scholar * Sehnal, D. et al. MOLE 2.0: advanced approach for analysis
of biomacromolecular channels. _J. Cheminformatics_ 5, 39 (2013). Article CAS Google Scholar Download references ACKNOWLEDGEMENTS We thank the Computing and Cryo-EM platforms of the
Tsinghua University Branch of the National Center for Protein Sciences (Beijing) for cryo-EM sample preparation and screening. We acknowledge computational support from Beijing Computational
Science Research Center (CSRC) and Beijing Three Axis Space Technology Co. Ltd. This work was supported by the National Program on Key Research Project of China (grant nos. 2020YFA0707500,
2017YFC0840300, 2018YFA0507200 and 2018YFE0200402), National Natural Science Foundation of China (31971126, 31770309, 81520108019 and 31670731) and Nature Science Foundation of Hubei
Province (2019CFB790). AUTHOR INFORMATION Author notes * These authors contributed equally: Panpan Wang, Lu Liu, Aijun Liu. AUTHORS AND AFFILIATIONS * MOE Key Laboratory of Protein Science
and Collaborative Innovation Center of Biotherapy, School of Medicine, Tsinghua University, Beijing, China Panpan Wang, Lu Liu, Aijun Liu, Liming Yan, Zihe Rao & Zhiyong Lou * School of
Life Sciences, Tsinghua University, Beijing, China Panpan Wang, Lu Liu, Aijun Liu, Mingxu Hu & Zihe Rao * State Key Laboratory of Medicinal Chemical Biology, College of Life Science,
Nankai University, Tianjin, China Yong He, Yu Guo & Zihe Rao * Drug Discovery Center for Infectious Disease, College of Pharmacy, Nankai University, Tianjin, China Yong He, Yu Guo, Fei
Deng & Zihe Rao * State Key Laboratory of Virology and National Virus Resource Center, Wuhan Institute of Virology, Center for Biosafety Mega-Science, Chinese Academy of Sciences, Hubei,
China Shu Shen & Fei Deng * Complex Systems Division, Beijing Computational Science Research Center, Beijing, China Haiguang Liu * Department of Biology, Southern University of Science
and Technology, Shenzhen, China Chuang Liu & Peiyi Wang * 302 Hospital, Peking University, Beijing, China Yinying Lu * National Laboratory of Macromolecules, Institute of Biophysics,
Chinese Academy of Sciences, Beijing, China Zihe Rao Authors * Panpan Wang View author publications You can also search for this author inPubMed Google Scholar * Lu Liu View author
publications You can also search for this author inPubMed Google Scholar * Aijun Liu View author publications You can also search for this author inPubMed Google Scholar * Liming Yan View
author publications You can also search for this author inPubMed Google Scholar * Yong He View author publications You can also search for this author inPubMed Google Scholar * Shu Shen View
author publications You can also search for this author inPubMed Google Scholar * Mingxu Hu View author publications You can also search for this author inPubMed Google Scholar * Yu Guo
View author publications You can also search for this author inPubMed Google Scholar * Haiguang Liu View author publications You can also search for this author inPubMed Google Scholar *
Chuang Liu View author publications You can also search for this author inPubMed Google Scholar * Yinying Lu View author publications You can also search for this author inPubMed Google
Scholar * Peiyi Wang View author publications You can also search for this author inPubMed Google Scholar * Fei Deng View author publications You can also search for this author inPubMed
Google Scholar * Zihe Rao View author publications You can also search for this author inPubMed Google Scholar * Zhiyong Lou View author publications You can also search for this author
inPubMed Google Scholar CONTRIBUTIONS Z.L., Z.R. and F.D. conceived of the project and designed the experiments. Panpan Wang, L.L., L.Y., Y.H., S.S. and Y.G. expressed and purified the
proteins. Panpan Wang, L.L. and A.L. prepared the cryo-EM sample. Panpan Wang, A.L., M.H., H.L., C.L. and Peiyi Wang collected and processed the cryo-EM data and built the model. Panpan Wang
and Y.L. performed the surface plasmon resonance experiments. Z.L. wrote the manuscript. All authors discussed the experiments and read and approved the manuscript. CORRESPONDING AUTHORS
Correspondence to Fei Deng, Zihe Rao or Zhiyong Lou. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer
Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. EXTENDED DATA EXTENDED DATA FIG. 1 CRYO-EM RECONSTRUCTION. A, Raw image of
SFTSV-L particles in vitreous ice recorded at defocus values of -1.5 to -2.5 μm. Scale bar, 100 nm. B, Power spectrum of the image shown in (A), with plot of the rotationally averaged
intensity versus resolution. White circle indicates the spatial frequency corresponding to 3.0 Å resolution. C, Representative class averages. The edge of each square is 238 Å. D, Overview
of SFTSV-L reconstruction is shown in the left panel. The close-up view of a representative region in the RdRp region is shown in the right panel. Density is shown as gray mesh; the
polypeptide of the refined model is displayed as colored sticks (carbons, green; nitrogen, blue; oxygen, red). E, Fourier shell correlation (FSC) of the final 3D reconstruction following
gold standard refinement. FSC curves are plotted before (red) and after (green) masking in addition to post-correction (blue), accounting for the effect of the mask using phase
randomization. F, Local resolution estimation was performed with Relion 2.11. G, The 3DFSC sphericity was analyzed with 3DFSC2. H, The data processing scheme used to obtain the final map.
EXTENDED DATA FIG. 2 SECONDARY STRUCTURE DIAGRAM OF SFTSV-L. Secondary structure elements along the SFTSV-L sequence are shown as cylinders and arrows for ɑ-helices and β-strands,
respectively. Domains are colored as in Fig. 1. EXTENDED DATA FIG. 3 THE LARIAT DOMAIN. A-C The lariat domain is represented as a cartoon diagram in red covered by cryo-EM density in three
views. Other domains of SFTSV-L are covered by molecular surface with the same color as in Fig. 1. D, A close-up view of the lariat domain covered by the cryo-EM density. Residues for
tracing the direction of the polypeptide are indicated by dots with labels. EXTENDED DATA FIG. 4 THE ENDON DOMAIN. A, The contacts between SFTSV-L endoN domain and other domains. The
structure of SFTSV-L is displayed with the same color scheme in Fig. 1. The endoN domain is covered by a molecular surface, and other domains are shown as cartoon diagrams. The back view and
the side view of the endoN domain are shown in the right panels. The interdomain interfaces on the endoN domain are indicated by the same color as each of the interacting domains, including
the RdRp core, the PB2-N-like domain, the arm domain and the blocker motif, the CBD domain and the lariat domain. B, The orientation of the endoN domains in SFTSV-L and LACV-L. The
structures of SFTSV-L and LACV-L 1750 are aligned according to their RdRp domains. The endoN domains of SFTSV-L and LACV-L are covered by molecular surface in forest green and smudge green,
respectively. The active residues of each endoN domain are highlighted in orange. The polypeptide of other domains in SFTSV-L was represented as cartoon diagram. The cap-binding pocket in
the CBD domain is highlighted by a red dotted circle. The distance from the cap-binding pocket to the position of the active center of LACV-L endoN domain is approximately 80 Å. EXTENDED
DATA FIG. 5 ORIENTATION OF THE CBD DOMAIN AND THE BLOCKER MOTIF. This figure is related to Fig. 2. The structures of SFTSV-L A, FluA-vRNA PA-PB1-PB2 complex B, FluB1-vRNA PA-PB1-PB2 complex
C, FluB-cRNA PA-PB1-PB2 complex D, and apo FluC PA-PB1-PB2 complex E, are aligned according to their CBD domains and are shown in the same orientation in the upper panels. Because the CBD
domains from these structures are located different site from their own RdRp cores, other domains of these structures are in significantly distinct positions. In the lower panels, the CBD
domain and its interacting fragments in these structures are shown in detail. The CBD domains are covered by molecular surface, and the other fragments are shown as cartoon diagrams. The
residues constituting the cap-binding pockets are highlighted in red and the PB2 cap-627 linker3 is in blue. Note, the orientation of the detailed illustrations for FluB1 (C), FluB (D) and
FluC (E) has a ~90° clockwise rotation for clear presentation. The residues occupy the cap-binding pockets of SFTSV-L and apo FluC PA-PB1-PB2 complex are displayed as spheres. EXTENDED DATA
FIG. 6 COMPARISON OF SUBDOMAINS IN THE RDRP REGION. The structures of the RdRp region from SFTSV-L, LACV-L and IAV PA-PB1-PB2 complex are in the same orientation and color scheme as in Fig.
4b. The PA-C-like (or PA-C) domain, the RdRp core, and the PB2-N-like (or PB2-N) domain are shown separately in the right, middle and left panels. EXTENDED DATA FIG. 7 COMPARISON OF THE
PRIMING LOOP. A-D. The subdomains of the RdRp core and the PA-C-like domain of SFTSV-L (A), LACV-L (PDB code: 5AMR) (C) and FluA PA-PB1-PB2 (PDB code: 4WSB) (D) are covered by a molecular
surface with the same color scheme as that in Fig. 4b. The priming loops in three structures are shown as white cartoons. The priming loop in SFTSV-L is enlarged and is covered with cryo-EM
density in (B). The RdRp catalytic centers of the three polymerases are highlighted by the white circles. E, Density of motif F. The RdRp motifs A, B, D, F, H of SFTSV-L are shown as cartoon
diagram with the same color scheme as that of Fig. 5a. Motif F is covered by the cryo-EM density. EXTENDED DATA FIG. 8 THE ACTIVE SITE OF THE RDRP DOMAIN. A, The details of the RdRp motifs
of SFTSV-L. This figure is generally the same as Fig. 5a, with one exception that the cation, assumed to be a magnesium ion, is shown as a red sphere. In contrast, two catalytic manganese
ions from the polio virus elongation complex structure4 are shown as black spheres. B, The bound cation and surrounding residues are covered by cryo-EM densities. The bonds formed between
SFTSV-L polypeptide and the cation are labeled with distance. C, The electrostatic surface potential of SFTSV-L RdRp tunnels. The positive surface is colored blue, the negative surface, red,
with limits ±70 kbT/ec. The regions belong to the NTP entry, template entry, template exit and product exit tunnles are indicated out by dashed lines. D, The entrance of NTP entry tunnel,
and E, the entrance of template entry tunnel. Colors are as in Fig. 4. Superposition of LACV-L 1750 (5AMR) structure shows the position of the 5' vRNA (gray cartoon) in (B). The
approximate positions of NTP and template entry tunnels are bordered by dashed lines. EXTENDED DATA FIG. 9 IMPACT OF THE ENDON DOMAIN ON THE PRODUCT EXIT TUNNEL. This figure is related to
Fig. 5b. A, Same as Fig. 5b, the tunnels for template entry, NTP entry, product exit, and template exit of SFVSV-L are indicated with violet, salmon red, orange and green labels in (A). B, A
model without the endoN domain and residues 206-217 in the linker region was generated, and its tunnels were calculated and shown as the same color scheme in (A). The bottom views of the
product exit tunnel in SFTSV-L C, or the model without the endoN domain and linker residues D, is shown. EXTENDED DATA FIG. 10 PROPOSED MODELS FOR TRANSCRIPTION INITIATION AND CO-FACTOR
BINDING. A, A proposed speculative model of SFTSV transcription initiation. SFTSV-L is schematically represented, with the cap-binding pocket (red hollow stars) in the CBD domain, the RdRp
catalytic center (deep red dots) and the active center of the endoN domain (orange stars) as marked. (1) During the early event of the cap-snatching process, the lariat domain loses its
interaction with the RdRp domain to allow the L protein to transform into a fully open state. Meanwhile, the endoN domain is pulled out from the central position of the SFTSV-L, and the arm
domain shifts to induce the removal of the Arg-finger in the block motif from the cap-binding pocket of the CBD domain. (2) After the host mRNA is captured, the endoN domain employs an ~180°
rotation along its long axis, thus causing its active center to face the nucleotide chain of host mRNA, and cleaves it by using two cations as cofactors chelated by the conserved
H...PD...D/E...K motif. If the structures of LACV-L 1750 and SFTSV-L are aligned using their RdRp cores as the reference, the cap-binding pocket in the SFTSV-L CBD domain will have a
distance of ~80 Å relative to the position of the LACV-L endoN domain active center (Supplementary Fig. 5). We reason the lack of C-terminal parts in the structure of LACV-L 1750 abolishes
the extensive interdomain interactions to maintain the compact structure and allow its endoN domain to be in a fully open state. (3) Then, the CBD anchors the snatched cap structure with
additional nucleotides and transfers the 3' end nucleotide chain to the RdRp catalytic cavity. (4) The transcription of virus mRNA is thus elongated with the guidance by vRNA as the
template, and the products will be released from the product exit tunnel. In this stage, the endoN domain is likely to stay out of the center of the L protein, allowing the product exit
tunnel to be in a fully open state. Meanwhile, the lariat motif will go back to its initial state. B, The structures of SFTSV-L and RSV-L-P complex5 are aligned by their RdRp core and shown
in the same orientation. The colors for SFTSV-L domains are the same as those in Fig. 1. The RdRp and the PRNTase domains of RSV-L are shown in pale cyan and cyan, respectively. The P
proteins bound to RSV-L is shown as cartoon diagram, and its structurally equivalent position in SFTSV-L is framed out. SUPPLEMENTARY INFORMATION SUPPLEMENTARY INFORMATION Supplementary Fig.
1 and Table 1. REPORTING SUMMARY RIGHTS AND PERMISSIONS Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Wang, P., Liu, L., Liu, A. _et al._ Structure of severe fever with
thrombocytopenia syndrome virus L protein elucidates the mechanisms of viral transcription initiation. _Nat Microbiol_ 5, 864–871 (2020). https://doi.org/10.1038/s41564-020-0712-2 Download
citation * Received: 25 November 2019 * Accepted: 19 March 2020 * Published: 27 April 2020 * Issue Date: 02 June 2020 * DOI: https://doi.org/10.1038/s41564-020-0712-2 SHARE THIS ARTICLE
Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided
by the Springer Nature SharedIt content-sharing initiative
Trending News
Environmental news, opinion and analysis from Guardian US | The GuardianEnvironmentHideCub found alone in US woods now being raised by wildlife staff in bear costumesTwo-month-old black bear, ...
Mehbooba hopes people come out in good numbers on may 25 to vote for pdpMehbooba hopes people come out in good numbers on May 25 to vote for PDP ISHRAT BHAT May 22, 2024 2:56 pm No Comments Vi...
Bitcoin hits record as stocks, bonds retreat amid market uncertaintyThe price of Bitcoin broke its record on Wednesday, climbing to just shy of $110,000 to eclipse the previous high from J...
CES 2016 : BMW i8 Mirrorless ConceptLe 06/01/2016 à 01:00, modifié le 14/02/2019 à 14:27 CLIQUEZ ICI POUR REVENIR A L'ARTICLE BMW I8 MIRRORLESS CONCEPTSur l...
Taylor swift finally airs reputation (taylor's version) track for iconic momentLOOK WHAT YOU MADE ME DO (TAYLOR'S VERSION), WHICH IS SET TO FEATURE ON THE ALBUM REPUTATION (TAYLOR'S VERSION...
Latests News
Structure of severe fever with thrombocytopenia syndrome virus l protein elucidates the mechanisms of viral transcription initiationABSTRACT Segmented negative-sense RNA viruses (sNSRVs) encode a single-polypeptide polymerase (L protein) or a heterotri...
How the pfizer covid vaccine gets from the freezer into your armFrom Monday, people at high risk of COVID-19 will be lining up to receive the Pfizer vaccine, marking the start of Austr...
Consortium releases online practice testsThe Smarter Balanced Assessment Consortium, one of two major groups of states designing tests matched to the Common Core...
Snowboarder shaun white withdraws from slopestyle eventSnowboarder Shaun White has announced that he is dropping out of the slopestyle event at the Sochi Olympics. The event i...
The expression of p-glycoprotein is causally related to a less aggressive phenotype in human osteosarcoma cellsABSTRACT The relationship between P-glycoprotein expression and malignancy is controversial. We have recently found that...