Role of rho gtpases in inflammatory bowel disease
Role of rho gtpases in inflammatory bowel disease"
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:
ABSTRACT Rat sarcoma virus homolog (Rho) guanosine triphosphatases (GTPases) function as “molecular switch” in cellular signaling regulation processes and are associated with the
pathogenesis of inflammatory bowel disease (IBD). This chronic intestinal tract inflammation primarily encompasses two diseases: Crohn’s disease and ulcerative colitis. The pathogenesis of
IBD is complex and considered to include four main factors and their interactions: genetics, intestinal microbiota, immune system, and environment. Recently, several novel pathogenic
components have been identified. In addition, potential therapies for IBD targeting Rho GTPases have emerged and proven to be clinically effective. This review mainly focuses on Rho GTPases
and their possible mechanisms in IBD pathogenesis. The therapeutic possibility of Rho GTPases is also discussed. SIMILAR CONTENT BEING VIEWED BY OTHERS THE ROLE OF THE HIPPO PATHWAY IN THE
PATHOGENESIS OF INFLAMMATORY BOWEL DISEASE Article Open access 12 January 2021 RECENT DEVELOPMENTS ON BMPS AND THEIR ANTAGONISTS IN INFLAMMATORY BOWEL DISEASES Article Open access 01 July
2023 HARNESSING MURINE MODELS OF CROHN’S DISEASE ILEITIS TO ADVANCE CONCEPTS OF PATHOPHYSIOLOGY AND TREATMENT Article 27 July 2021 FACTS * Rho GTPases are vital in numerous cellular events
and associated with IBD pathogenesis. * Rho GTPases participate in traditionally believed pathogenic factors of IBD. * Roles of Rho GTPases in several novel pathogenic factors of IBD are
also proposed. OPEN QUESTIONS * Exactly what roles Rho GTPases play in IBD pathogenesis and what are the specific mechanisms? * How to optimize the experimental design to explore the
connections between Rho GTPases and IBD? * Can we invent more secure and effective methods for IBD treatment by targeting Rho GTPases? INTRODUCTION Characterized by progressive and chronic
relapsing intestinal inflammation, inflammatory bowel disease (IBD) is likely to occur early in life and is unfortunately incurable [1]. Despite having an unclear pathogenesis, studies have
indicated the involvement of genetic variants, environmental changes, abnormal intestinal microbiota, and immune response dysregulation, and the interactions between these factors ultimately
leads to the onset of IBD [2]. In recent years, many novel pathogenic factors have been identified that provide new insights into the disease [2]. Rat sarcoma virus (Ras) homolog (Rho)
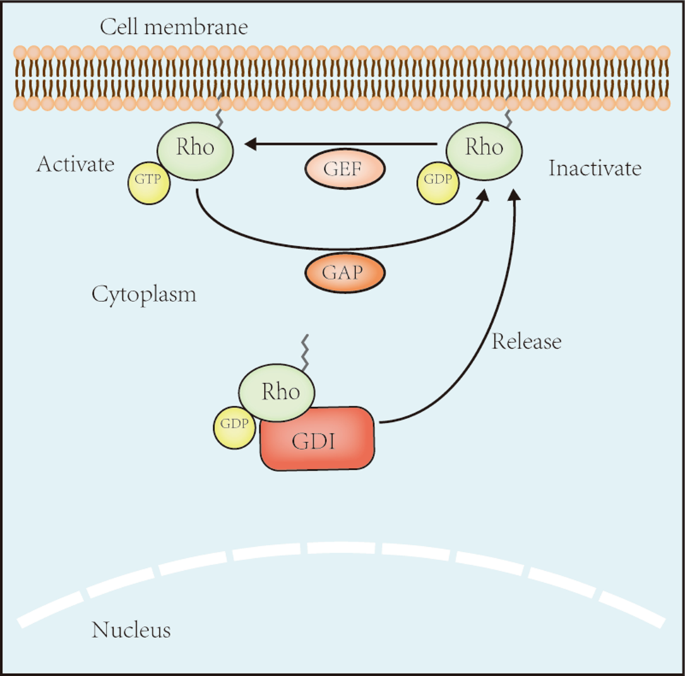
guanosine triphosphatases (GTPases), a subgroup of the Ras superfamily, are guanosine triphosphate (GTP)-bound proteins [3]. The regulation of Rho GTPase activity depends on guanine
nucleotide exchange factors (GEFs), GTPase-activating proteins (GAPs), and guanine nucleotide dissociation inhibitors (GDIs). GEFs enhance the switch of Rho GTPases from guanosine
diphosphate (GDP)-bound forms to GTP-bound forms to activate them, whereas GAPs and GDIs inactivate them by increasing bound GTP hydrolysis and binding directly to them, respectively [4]
(Fig. 1). Rho GTPases are vital in the cytoskeleton regulation and gene expression [3, 5]. Recently, studies have found that Rho GTPases are associated with numerous cellular events,
therefore being important in chronic diseases such as IBD. Within the Rho GTPase family, cell division cycle 42 (Cdc42), Ras-related C3 botulinum toxin substrate 1/2 (Rac1/2), and Rho family
member A (RhoA) are the most thoroughly studied proteins. In this review, the functions of Rho GTPases in the pathogenesis of IBD and their possible mechanisms are summarized. The
therapeutic strategies associated with Rho GTPases that may be beneficial to IBD patients and their clinical applied prospect are also discussed. GENETICS In recent years, over 200 loci have
been found to be implicated in IBD. Among these loci, most increase the risk of both IBD subtypes, whereas several are unique to either CD or UC [6]. Rho GTPases are involved in this
pathogenic process (Table 1). GENES ENCODING RHO GTPASES Rac2 deficiency is associated with worsening colitis in mice treated with _Citrobacter rodentium_, implying a possible effect of this
protein on human intestinal diseases development [7]. This hypothesis is supported by the fact that several _Rac2_ single nucleotide polymorphisms (SNPs) may function in the predisposition
and susceptibility to CD [8, 9]. Interestingly, whether the association between Rac2 and IBD is based on the function of Rac2 in the NADPH complex is controversial [7, 8]. The _Rac1_ SNPs
rs10951982 and rs4720672 are associated with UC [10], and another _Rac1_ SNP, rs34932801, was found to lead to poorer thiopurine treatment effect in adult CD patients [11]; however, the
association between _Rac1_ SNPs and UC was not observed in another study [12]. In addition, mice with a genetic deletion of _RhoA_ in intestinal epithelial cells (IECs) exhibited spontaneous
chronic intestinal inflammation, and RhoA signaling was found to be downregulated in CD patients [13]. OTHER GENES ASSOCIATED WITH RHO GTPASES MYO9B Genetic variations in myosin IXB
(_Myo9B_), a Rho GAP encoding gene, are associated with IBD [14, 15]. This association might be partly explained by the dysfunction of Myo9B, leading to an abnormal level of active Rho,
subsequently influencing intestinal epithelial wound healing and disrupting the formation of tight junctions (TJs) [16]. PGGT1B Protein geranylgeranyltransferase type-I (GGTase-I) transfers
a 20-carbon geranylgeranyl lipid to substrate proteins, including Rho GTPases [17]. This process is a post-translational modification of Rho GTPases called prenylation. Genetic deletion of
_Pggt1b_, a gene encoding GGTase-I, in murine IECs results in epithelial injuries and spontaneous gut inflammation, which can be significantly ameliorated by activating RhoA signaling [13].
Loss of _Pggt1b_ in the T cells of mice also causes spontaneous colitis via impaired RhoA signaling, and the intestinal T cells of IBD patients have lower _Pggt1b_ level [18]. ATG16L1 The
SNP T300 of autophagy-related protein 16-like protein 1 (_ATG16L1_) contributes to CD. In mice, defective autophagy stimulates Rac1 to restrain dendritic cells (DCs) migration [19] and
inhibits RhoA activity to affect IECs migration to restore the intestinal mucosa [20]. NCF2 The neutrophil cytosolic factor 2 (_NCF2_) gene is considered a very early onset IBD-specific
susceptibility gene. A missense variant of this gene causes abnormal binding of its p67phox product to Rac2, resulting in a damaged function of oxidase and thus contributing to the
development of IBD [8]. TTC7A Lemoine et al. [21] identified biallelic missense mutations in tetratricopeptide repeat domain 7 A (_TTC7A_) that inappropriately activated RhoA signaling in
IBD patients with an immune deficiency. TAGAP A study showed that T cell activation of the Rho GTPase-activating protein (_TAGAP_) SNP rs212388 negatively correlated with the severity of
anal disease in CD, and hypothesized that the protective effect may be attributed to impaired Rho GTPase inactivation [22]. This supposition requires more studies to authenticate. INTESTINAL
MICROBIOTA Studies have shown for decades that the intestinal microbiota is essential to IBD development [23]. Intestinal homeostasis partly depends on the mutual influence of intestinal
microbiota and the immune system [24] and Rho GTPases are involved in the interaction. For example, segmented filamentous bacteria (SFP), which are commensal gut bacteria in several animal
species, induce the differentiation of T helper 17 (Th17) cells in the mouse intestine, which requires Cdc42-dependent endocytosis [25]. Compared to normal person, IBD patients intestinal
microbiota communities are markedly different, with decreased abundance of Firmicutes and _Saccharomyces cerevisiae_ and increased abundance of Proteobacteria (especially Enterobacteriaceae)
and _Candida albicans_ [26, 27]. Additionally, the microorganisms expanded in IBD patients can experimentally induce colitis, whereas those contracted are able to ameliorate it [28].
_Escherichia coli_, especially adherent-invasive _E. coli_, participate in IBD, and adherent-invasive _E. coli_ strains share similarities with extraintestinal pathogenic _E_. coli such as
uropathogenic _E. coli_ [29]. Cytotoxic necrotizing factor 1, a protein produced by uropathogenic _E. coli_, modifies Rac2 to restrain it in an active state. Active Rac2 then induces
receptor-interacting proteins 1 and 2 (RIP1 and RIP2) to trigger nuclear factor-kappa B (NF-κB) and interferon regulatory factor (IRF) pathways, driving a protective immune response in
mammalian cells [30]. Although the connection between enteropathogenic and enterohemorrhagic _E. coli_ and IBD has seldom been studied, it has been disclosed that their pathogenesis involves
the activation of downstream substrates Cdc42, Rac1, and RhoA as RhoGEFs [31]. Interestingly, with a similar mechanism, _C. rodentium_ infects colon cells, inducing IBD-linked inflammation
in mice [32]. Moreover, _Salmonella_ spp. (family Enterobacteriaceae) produce the pro-inflammatory protein SopE to activate Rac1 and Cdc42, subsequently inducing nucleotide oligomerization
domain-containing protein 1 (NOD1)/RIP2/NF-κB signaling, causing an inflammatory response in host cells [33, 34]. Taken together, these studies suggest that Rho GTPases participate in
inflammation induced by expanded Enterobacteriaceae in the gut mucosa, thus contributing to the onset of IBD. Additionally, _Clostridium difficile_ influences the deterioration and
recurrence of IBD [35] probably because its enterotoxins inactivate Rho, Rac, and Cdc42 [ref. [36]]; however, whether this bacterium is associated with IBD development remains unknown.
Accordingly, the role and mechanism of action of aforementioned bacteria in IBD require further research. IMMUNE SYSTEM THE INNATE IMMUNE SYSTEM The innate immune system includes
non-classical (intestinal epithelial barrier) and classical factors (DCs, macrophages, monocytes, and neutrophils). Dysregulation of these protective components by Rho GTPases is associated
with IBD [2, 37] (Table 2). INTESTINAL EPITHELIAL BARRIER Dysregulation of the intestinal epithelial barrier, including abnormal mucous layer [38], damaged structures and functions of the
IECs [36], and decreased expression of antimicrobial peptides [39, 40], have been reported in IBD. Increased TJs permeability, altered cytoskeletal rearrangement, and abnormal cell death in
IECs are strongly associated with IBD [13] and Rho GTPases influence IECs through all these aspects. Interestingly, both the overactivation and inactivation of Rho GTPases damage TJs by
directly affecting TJ proteins and indirectly affecting the F-actin cytoskeleton intimately associated with TJs [36, 41]. Reduction of cortactin, an actin-binding protein reduced in IBD
patients, upregulates RhoA/Rho-associated protein kinase (ROCK) signaling to phosphorylate myosin light chain, which contracts actomyosin leading to altered TJs and increased epithelial
permeability [42]. As mentioned above, MYO9B deficiency probably results in the abnormal activation of Rho to interrupt the formation of TJs [16]. Reduced RhoB levels are also associated
with impaired TJs that contribute to UC [43] and increased RhoB expression combined with decreased Cdc42 expression ameliorates dextran sulfate sodium (DSS)-induced increased intestinal
permeability and IEC apoptosis [44]. Impaired RhoA prenylation and signaling in IECs found in IBD patients drive altered cytoskeletal rearrangements [13]. The cytokines interferon-gamma
(IFN-γ) and tumor necrosis factor-alpha (TNF-α) are increased in IBD patients [45], inducing TJ proteins endocytosis and IECs apoptosis through the RhoA/ROCK pathway and the activation of
Rac1 triggering the c-Jun N-terminal kinase (JNK) pathway, respectively [46, 47]. _Clostridium difficile_ toxins induce IEC apoptosis in a Rho GTPase-dependent manner [48, 49]. Defective
mucosal healing is also a characteristic of IBD. RhoA deficiency in IECs is related to CD, with defective IECs migration and intestinal injuries repairment [20], while Rac1 inhibition
suppresses IECs migration and inhibits the recovery of colonic wounds in DSS-induced mice [50]. Cdc42 also contributed to mucosal healing in DSS-treated mice [51]. Compared to UC, reduced
expression of inducible β- and α-defensins secreted by Paneth cells has been observed in patients with colonic CD [39, 40]. Studies also identify that α-defensins inhibit _Clostridium
difficile_ toxins by protecting Rac1 from glucosylating [52, 53], while inducible β-defensins improve intestinal wound healing in mice by activating RhoA/ROCK signaling [54]. DCS AND
MACROPHAGES Dendritic cells and macrophages play both similar and different roles in maintaining intestinal homeostasis and inducing an immune response. DCs mainly function as
antigen-presenting cells (APCs), as they uptake and present antigens to regulate immune responses, and participate in lymphocyte gut homing [55]. Although macrophages are also considered
APCs, their major functions are engulfing and clearing pathogens and apoptotic cells to resolve inflammation [56]. The onset of IBD can be partly attributed to DCs and macrophages incorrect
recognition of commensal microorganisms, their abnormal activation and migration, and the subsequent imbalance between immune tolerance and activation [55, 57]. Impaired clearance of
microorganisms and apoptotic cells and inflammatory resolution have also reported at the onset of IBD [58]. Endocytosis is associated with antigen uptake and engulfing, which have been shown
to depend on Cdc42 and Rac regulation in DCs [59, 60]. In macrophages, this process varies according to the receptor. Cdc42 and Rac1 are recruited to the plasma membrane where Fc gamma
receptor (FcγR) is clustered, which then recruits Wiskott-Aldrich syndrome protein to activate the actin-related protein 2/3 complex. The latter activates p21-activated kinase (PAK) and
phosphatidylinositol-4-phosphate 5-kinase to regulate FcγR-mediated endocytosis in macrophages [61, 62]. Rac2, RhoG, and RhoC may also participate in this process [63, 64]. Studies have
demonstrated that Rac1 promotes toll-like receptor (TLR) 4-mediated bacterial phagocytosis [65] while RhoA and RhoG are required for complement receptor 3-mediated phagocytosis [66].
Furthermore, apoptotic cell engulfing mediated by integrin is enhanced by Rac1, Cdc42, and RhoG, and inhibited by RhoA [67, 68]. DCs require Rac, Cdc42, and Rho involvement to accomplish
migration and antigen presentation; the former requires the dynamic regulation and cooperation of Rho GTPases. Rac1/2 and Cdc42 are indispensable in forming lamellipodia and filopodia at the
leading edge, respectively, while RhoA controls contractility at the cell rear and migration [69, 70]. In macrophages, the maturation of phagosomes is promoted by Rho/ezrin-radixin-moesin
(ERM) protein [71] and macrophages then pretend to exert a microbicidal function. Rac2 is an essential component of the NADPH complex [8] but Rac1 has also been shown to be involved. After
phagocytosis, Rac1 is activated and recruited to the plasma membrane, causing the NADPH complex activation and reactive oxygen species generation to remove microorganisms[72, 73].
Furthermore, macrophage complement is a process that includes monocyte adhesion, migration, and differentiation. RhoA and Cdc42 are required for monocyte adhesion, whereas migration requires
Rac, RhoA, and Cdc42 participation [74]. The migration mechanism is similar to that aforementioned. Another study found that RhoA functioned in both the front and back of cells,
contributing to leading edge construction and governing turning while stimulating actomyosin contractility, respectively [75]. In brief, these studies imply that Rho GTPases may contribute
to abnormal DCs and macrophage functions induce IBD pathogenesis; however, direct evidence is lacking to confirm the connection between them. NEUTROPHILS Abnormal activation, recruitment,
and function of neutrophils have been identified in IBD pathogenesis [76, 77]. In neutrophils, Rho GTPases play roles similar to that in DCs and macrophages, i.e., they regulate migration,
phagocytosis, and microbe clearing [78, 79], in addition to the degranulation process [80]. A few findings directly demonstrate that dysfunction of Rho GTPases in neutrophils contributes to
IBD. Rac1 deficiency in mouse neutrophils ameliorated DSS-induced colitis, probably by inhibiting neutrophil recruitment and migration to the inflammation site [10]. Failure of Rac2 to bind
the NADPH complex in neutrophils results in susceptibility to CD [8]. THE ADAPTIVE IMMUNE SYSTEM The adaptive immune system includes T cell-induced cellular immunity and B cell-induced
humoral immunity, and the involvement of the former in IBD has been extensively explored [2]. Rho GTPases are crucial for regulating lymphocyte biology [81]. T CELLS Abnormal T cell
activation, accumulation, differentiation and inappropriate apoptosis are considered IBD immunopathogenesis [2]. Rho GTPases are crucial in adaptive immune response because of their wide
control of T cell basic functions (Fig. 2). MIGRATION Migration of T cells is a sophisticated process. The mechanism of this event is similar, requiring cooperation between Rac, Cdc42, and
RhoA [75, 82]. In addition, there is accumulating evidence that Rho GTPases contribute to T cell recruitment. C-X-C motif chemokine ligand 12 (CXCL12) stimulates the Rac1 guanine nucleotide
exchange factor (Vav1)/Rac1 pathway to upregulate integrin α4β1 expression to promote T cell adhesion [83], a process that might be RhoA-dependent [84]. Without Rac2, T cells show defective
response to CXCL12 and chemokine ligands 19 and 21 and are less efficient at homing to lymph nodes [85]. Once T cells are effectively activated, Rac and RhoA start to inhibit migration. The
function of Rac here depends at least partly on the dephosphorylation of ERM proteins [82, 83]. RhoH also negatively regulates T-cell migration [86]. In IBD patients, T cells isolated from
gut tissues showed decreased expression of _Pggt1b_, which led to dysfunction of RhoA in IECs and T cells in mice, promoting CD4 + T cell accumulating at the intestine and aggravating
colitis [13, 18]. ACTIVATION The interaction between T cells and APCs, an essential process for triggering T cell activity, is induced by T cell antigen receptor (TCR) and co-stimulatory
molecules, followed by an immunological synapse formation, where active Rac1, Cdc42, and RhoA are observed [87]. Additionally, Rac enhances the connection between T cells and APCs by
participating in the dephosphorylation of ERM proteins [88], while inactive Cdc42, Rac, and RhoA impair the activity of co-stimulatory molecules [89, 90]. The upcoming signal transduction
process also requires Rho GTPases involvement. The activation of JNKs and transcription factors activator protein-1, nuclear factor of activated T cells, and NF-κB is associated with
Rac1[refs. [91, 92]]. Rac1 also influences Ca2+ influx into T cells [93], an event involved in the maintenance of this interaction [94] and in cytoskeleton reorganization during signal
transduction [94, 95]. The latter probably depends on the phosphoinositide 3 kinase (PI3K)/Vav1/Rac or Rho pathway [95]. Inhibition of RhoA leads to damaged Ca2+ influx [96] and RhoA is
involved in T cell activation through the regulation of mitochondrial function [97]. Cdc42 deficiency results in the overactivation of murine T cells [98]. Moreover, RhoG and RhoH have been
reported to regulate the TCR signaling pathway, mainly as the suppressor and enhancer, respectively [86, 99]. DIFFERENTIATION After activation, T cells differentiate into various subtypes
that perform different functions. Generally, CD was considered a Th1 condition and UC an atypical Th2 condition. With the observation of other subsets engagement, such as Th17 and regulatory
T cells (Tregs), this view has changed [2]. As mentioned above, Cdc42 is required for SFB-induced Th17 differentiation in mice [25]. Deficiency of Cdc42 promotes the differentiation and
pathogenicity of Th17 and disrupts Tregs differentiation and stability, aggravating DSS-induced colitis [100]. Deficiency of Cdc42 in mice also interrupts Th2 differentiation [101]. RhoA
induces Th2 and Th17 differentiation, and the former occurs partly via defective T cell metabolism [97, 102]. However, another study suggested that decreased activation of RhoA, because of
its abnormal membrane location, contributes to a shift from Th1- to Th2-modulated immunity through the inhibition of p38 [103]. In Th1 cells, Rac2 triggers the NF-κB and p38 signaling
pathways to upregulate IFN-γ expression [104]. As for RhoH, a study found a remarkably higher differentiation level of Th1 than Th2 [105]. Additionally, decreased RhoB activity promotes
Tregs differentiation [106]. APOPTOSIS Cdc42 activates JNK and upregulates Fas and Fas ligand (FasL) expressions to induce T cell apoptosis in secondary lymphoid organs [107]. Cdc42 and Rac
are associated with both the induction and late stages of Fas/FasL-induced apoptosis but do not directly participate in it [108], and activated Rho/ROCK signaling is also involved in the
process by phosphorylating ERM proteins [109]. Interestingly, decreased RhoA activation has been reported to contribute to T-cell apoptosis [110]. Activated Rac1 with Vav1 restrains the
NF-κB and signal transducer and activator of transcription 3 (STAT3) signaling pathways to induce T cell apoptosis to alleviate clinical symptoms in IBD patients [111, 112]. CYTOKINES
Cytokines include interleukins (ILs), colony stimulating factors (CSFs), IFNs, TNFs, transforming growth factor-β family members, growth factors, and chemokines. The role of these molecules
has been gradually explored in IBD [113]. Cdc42 has been shown to downregulate IL-4, IL-10, TNF-α, and IFN-γ expression both in the colon tissues and serum of 2,4,6-trinitrobenzenesulfonic
acid (TNBS)-induced mouse models to regulate inflammation [114]. Conversely, TNF-α downregulated Cdc42, while CdC42 overexpression reversed the intestinal epithelial barrier damage due to
TNF-α in Caco2 cells [115]. RhoA/ROCK pathway activation stimulates the production of TNF-α and IL-1β, and there is a positive correlation between RhoA and TNF-α in the intestinal
inflammatory tissues of CD patients [116]. Inhibition of the RhoA/ROCK pathway also disrupted TGF-β-induced fibrosis in human [117]. RhoA dysfunction in T cells promoted IL-1β, IL-17A,
TNF-α, and IFN-γ expression in mice developing spontaneous colitis [18] and blocking RhoA activation reduced IL-2 production [96]. In addition, Horowitz et al. [118] identified that
TNF-α/lipopolysaccharide could activate the RhoA/ROCK pathway and contribute to IBD. Rac1 activity is upregulated by TGF-β in Caco2 cells to promote migration and epithelial restitution
[119]. Rac1 can also be activated by CSF, IL-6, IL-8, and TNF-α [120]. Inhibition of Rac1 decreases IL-8 level, thus regulating immunocyte migration to the intestinal tract to maintain local
remission [120, 121]. Rac1 also suppresses tryptophan-induced reduction of TNF-α, which damages intestinal barrier [122]. In DSS-treated mice, Rac1 deficiency in macrophages and neutrophils
results in lower levels of IL-1β, IL-12, and TNF-α [10] whereas a Rac1 inhibitor blocks engulfment and cell motility protein 1-mediated downregulation of IL-1α, IL-6, IL-12B, IL33, TNF-α,
and TGF-β to disrupt its protective function in DSS-induced mice [50]. Furthermore, Rac2 is associated with IFN-γ and IL-17A production, suggesting that impaired Rac2 function leads to IBD
development [7, 104], and Rac2 inhibition can be used to treat IBD patients [112]. ENVIRONMENT Smoking has been considered as a risk factor for CD but a protective factor for UC [123, 124].
Nicotine, a component of cigarettes, activates α7 nicotinic acetylcholine receptors (α7nAChRs) on plasmacytoid DCs to disrupt their migration into colonic mucosa. This protects cells from UC
via activated JAK2-STAT3 signaling and the subsequent degradation of Rac1 induced by non-apoptotic caspase-3 [ref. [124]]. However, whether the mechanism of smoking-induced CD involves Rho
GTPases remains unclear. The connections between Rho GTPases and other environmental factors contributing to the onset of IBD [124] have been scarcely explored. A study identified that the
activation of vitamin D receptor via the RhoA/Rho kinase pathway partly inhibited the inflammatory response in benign prostatic hyperplasia [125] and myocardial ischemia-reperfusion injury
[126]. In addition, vitamin D deficiency is associated with IBD [123], suggesting a possible involvement of Rho GTPases in vitamin D-deficiency-induced IBD. Therefore, there is an urgent
need to investigate this association. NOVEL PATHOGENIC COMPONENTS In addition to the aforementioned classical pathogenic factors, several novel components in the pathogenesis of IBD, such as
pattern recognition receptors (PRRs), non-coding RNAs (ncRNAs), angiogenesis, and fatty acids, have gradually drawn researchers’ attention [2, 127, 128]. Rho GTPases are also participants
in these pathogenic processes. PRRS Pattern recognition receptors, including receptor families such as TLRs and NOD-like receptors (NLRs), recognize pathogen-associated molecular patterns
and damage-associated molecular patterns to initiate an immune response and regulate other events [129]. TLRS TLRs are transmembrane proteins that participate in innate immunity. Upon
activation, TLRs trigger downstream signaling cascades to activate NF-κB, which results in pro-inflammatory mediators secretion to regulate adaptive immunity. The expression and function of
TLRs are irregular in IBD patients, suggesting that TLRs are both positively and negatively associated with IBD [130]. Rho GTPases take part in TLR-mediated immune response. In monocytes,
Rac1 and RhoA are vital in TLR2-induced NF-κB activation via a Rac1/PI3K/protein kinase B pathway [131] and at least partially through atypical protein kinase C [132]. Rac1 also participates
in the TLR1-mediated signaling pathway in epithelial cells [133]. Rac, RhoA, and Cdc42 are all activated after TLR4 is stimulated to promote uptake by macrophages [134, 135].
TLR7/9-mediated production of type I IFN requires Rac involvement in DCs [136]. In macrophages, following TLR activation, Rac acts as a positive regulator of phagocytosis and oxidative burst
because of the downregulation of TNF-α-induced protein 8-like 2 [ref. [137]] whereas RhoB enhances the expression of pro-inflammatory mediators by binding to major histocompatibility
complex class II [138]. In addition, Cdc42 is involved in TLR-activated DCs-T cells interactions [139]. NLRS NLRs are a group of cytoplasmic proteins whose downstream signaling pathways
mainly result in NF-κB and caspase-1 activation [140]. Mutations and dysfunction of NLRs are associated with IBD [140]. Rho GTPases are implicated in the functions exerted by NLRs. After
activation, NOD1/2 starts to form a complex (the nodosome) containing Rho GTPases, which is necessary for the activation of the complex itself and NF-κB eventually [34, 141]. Interestingly,
Rac1 negatively regulates the NOD2-mediated NF-κB activation and IL-8 induction [142]. In mouse macrophages, the NLR family pyrin domain containing protein 3 (NLRP3) inflammasome senses Rac2
activation and is then phosphorylated by p21-activated kinase 1/2, the substrate of Rac2, to induce the secretion of IL-1β, which contributes to microorganisms clearance [143]. Rho A is
also a positive regulator of NLRP3 in Caco2 cells [144]. Rac1 and RhoB are associated with the NLRP3 inflammasome as well [145, 146]. NCRNAS MicroRNAs (miRNAs, miR) are among the most
extensively studied ncRNAs, as they can silence gene expression by regulating messenger RNAs (mRNAs) translation or degradation [147]. Associations between miRNAs and IBD have been proposed
[148,149,150]. For instance, MiR-15a inhibits Cdc42 expression in pediatric IBD patients, resulting in a damaged intestinal epithelial barrier [116]. MiR-21 targeting and regulation of RhoB
expression, which is upregulated in UC, disrupts the intestinal epithelial barrier by damaging TJs [43]. Cdc42 is also regulated by miR-21 in the intestinal epithelial barrier regulation
[44]. MiR-31-3p alleviates colitis in IBD mouse models, possibly by targeting and negatively regulating RhoA expression, thus reducing the levels of several cytokines in colonic epithelial
cells [151]. Circular RNAs (circRNAs), another class of ncRNAs with covalently closed circles, interact with miRNAs and proteins to perform their functions [152]. The circRNA
homeodomain-interacting protein kinase 3 (circHIPK3) was recently identified as a sustainer of intestinal epithelium homeostasis and was reported to be decreased in patients with IBD [153].
CircHIPK3 is, at least partially, directly connected to and thus inhibits miR-29b availability to enhance the expression of Rac1 and Cdc42, promoting epithelial recovery in wounded IECs as
well as intestinal epithelium renewal in mice [153]. ANGIOGENESIS One crucial event in chronic inflammatory diseases is angiogenesis, a process enhanced in the intestinal inflamed site of
IBD patients, and vascular endothelial growth factor (VEGF) and IL-8 seem to act as inducers of this process [154]. Rho GTPases are implicated in this process, mainly in controlling cell
migration [155]. Activated Rac1, Cdc42, and Rho, probably via steroid receptor coactivator/focal adhesion kinase signaling, participate in VEGF-induced angiogenesis [156, 157]. Inhibition of
Rac1 and RhoA cause defected IL-8-induced endothelial cell migration [158]. Nevertheless, future studies should address the direct evidence of Rho GTPases as participants in angiogenesis,
contributing to disclose the molecular development of IBD through more mechanistic studies. FATTY ACIDS Whether the intake of fatty acids is beneficial or detrimental in IBD is still
controversial, as several studies have achieved different conclusions [127, 128]. Turk et al. [159] found that n-3 polyunsaturated fatty acids damaged epidermal growth factor
receptor-mediated Rac1 and Cdc42 activation to delay intestinal wound healing in DSS-treated mice. CLINICAL PROSPECT OF RHO GTPASES Emerging studies have shown that Rho GTPases act as
therapeutic targets in many diseases, including cancer [160], cardiovascular disease [161], and Alzheimer’s disease [162]. As Rho GTPases have a great influence on IBD pathogenesis, their
role in IBD treatment has been gradually explored (Table 3). CDC42 Antrum mucosal protein 18 (AMP-18), which is produced by antral mucosal epithelial cells [163], positively regulates
intestinal epithelial TJs stability and mucosa recovery [164]. Cdc42 is activated after treating with AMP-18 and contributes to the formation of TJs [165]. The anti-TNF antibody infliximab,
for example, has been proven to be effective in the induction and maintenance of clinical remission in IBD patients [166,167,168,169]. Recently, a study found that Cdc42 is upregulated in
patients with UC in response to infliximab [170], but the mechanism underlying this upregulation still requires further studies. RAC1 Nsc23766 is a Rac1-specific inhibitor that targets Rac1
binding and activation by RhoGEFs [171]. By moderately blocking Rac1 activation, Nsc23766 enhances phagocyte function in CD patients regarding the ability to clear the microorganisms [120].
Thiopurines is an effective drug to treat IBD [172] and 6-thioguanine triphosphate, one of the main metabolic products that exerts such function, directly binds to Rac1 to inhibit its
activation [173]. A clinical trail found that IBD patients response to thiopurine therapy had a lower Rac1 level or activity [174]. Inactivated Rac1 has distinct effects on various cells.
Inactivated Rac1 improves the defective migration of DCs in CD patients with the ATG16L1 mutation [19]. In monocytes, when Rac1 activity is inhibited within a certain range, phagocytic
function is enhanced [120] but in T cells this change results in a reduced immune response and increased T cell apoptosis due to the suppression of T cell-APC conjugation and impaired NF-κB
and STAT-3 signaling, respectively [111, 112]. In Caco2 cells, the migration of macrophages and neutrophils to the gut is suppressed by the downregulation of IL-8 expression [121]. In
addition, NK cells, whose role in IBD has been recently discovered, undergo enhanced induction of apoptosis mediated by caspase-9 [ref. [175]]. Overall, thiopurines seems to function as Rac1
inhibitors in IBD treatment. Another kind of clinical drug statins are also demonstrated to suppress Rac1 and RhoA [176] and alleviate CD patients’ inflammation [177]. Moreover, a few
clinical trials, working on estimating the efficacy and safety of statins in UC patients, are about to start. However, the activation of Rac1 also shows a possible benefit in IBD patients
because active Rac1 improves IECs migration and mucosal healing [50, 119]. RHOA Inhibitors of RhoA and its downstream effector ROCK are C3 exoenzyme [178] and Y-27632 [179], respectively.
Both C3 exoenzyme and Y-27632 disrupt arginase expression in human intestinal microvascular endothelial cells, which likely promotes intestinal microvascular endothelial function [118].
Y-27632 has been widely used in previous studies. By interrupting the RhoA/ROCK pathway, Y-27632 has been shown to improve TNBS-induced colitis in rat by inhibiting NF-κB activation [116],
alleviating DSS-induced intestinal inflammation by regulating cell function and cytokine secretion [180], and repress fibrogenesis by blocking RhoA/ROCK/MYLK/SRF signaling in colonic
myofibroblasts, representing a new therapy for intestinal fibrosis in IBD patients [117]. Furthermore, several substances have shown therapeutic potential by affecting RhoA/ROCK signaling.
Oxymatrine, a Chinese herb extract [181], and umbilical cord- and placenta-derived mesenchymal stem cells [182] suppress this pathway and thus alleviate intestinal inflammation in
DSS-treated mice [181] and inhibit intestinal fibrosis [182], respectively. However, blockade of signaling can also be detrimental. Partially hydrolyzed guar gum, a product of guar gum, was
observed to ameliorate colitis in mice [183]. Recently, a study has found that it improves wound healing in colonic epithelial cells, which is inhibited by Y-27632 [ref. [184]]. Thus,
RhoA/ROCK signaling pathway regulation might provide a new insight into IBD treatment. β-glucan, which upregulates the expression of Cdc42, Rac1, and RhoA, enhances cell migration and
proliferation, and ultimately promotes intestinal mucosal wound recovery, also represents a therapeutic potential for IBD via mucosal healing [185]. Although so far Rho GTPases targeted
drugs haven’t been widely applied and tested in clinic, many studies have proven that several IBD therapies influence the signal transduction pathway of Rho GTPases and their regulators are
effective in improving intestinal inflammation in animal experiments. Additionally, inhibition of Rho GTPases are used in other immunological disease treatments, such as rheumatoid
arthritis[186] and asthma [187]. Therefore, regulation of Rho GTPases might be clinically useful in treating IBD but still waits for further researches and larger clinical trials.
CONCLUSIONS Accumulating evidence has revealed that Rho GTPases are associated with IBD pathogenesis. According to the aforementioned studies, whether Rho GTPases have a beneficial or
detrimental effect on IBD development remains unclear. Studies of Rho GTPases in IBD conducted so far have been mostly based on static conditions using cells and/or animal models, and have
been limited by the deficiency of clinical samples. As numerous cell events depend on Rho GTPase dynamics, it is difficult to draw a connection between Rho GTPases and the development of
IBD. Nevertheless, therapies targeting Rho GTPases and their regulators are promising. Rho GTPase inhibitors have been widely studied and have been shown to be beneficial in alleviating
disease. In addition, the mechanism of thiopurine therapy, one of the most used treatments in the clinical setting, requires Rho GTPase inactivation. However, several studies pointed out
that Rho GTPase activation could also be a promising therapy. Together with the fact that extensive basic fundamental events require Rho GTPases involvement, general inhibition of Rho
GTPases might not only provide the desired therapeutic effect, but also be detrimental to the individual [188]. Additionally, thiopurine therapy is limited by its long-term treatment and
potential side effects. Thus, new drug discovery and fewer adverse reactions are future research directions for therapies targeting Rho GTPases. Furthermore, future studies should also focus
on other Rho GTPases that have been implicated in the onset of IBD. In summary, Rho GTPases are associated with the pathogenesis of IBD, and further clinical studies are required to confirm
and explore the underlying mechanisms. The Clinical Prospect of Rho GTPases is promising and require further studies on safety improvements and novel drug research development. DATA
AVAILABILITY All data included in this review are available upon request by contact with the corresponding author. REFERENCES * Agrawal M, Spencer EA, Colombel JF, Ungaro RC. Approach to the
Management of Recently Diagnosed Inflammatory Bowel Disease Patients: A User’s Guide for Adult and Pediatric Gastroenterologists[J/OL]. Gastroenterology. 2021;161:47–65. Article Google
Scholar * De Souza HSP, Fiocchi C. Immunopathogenesis of IBD: current state of the art[J/OL]. Nat Rev Gastroenterol Hepatol. 2016;13:13–27. Article Google Scholar * Jaffe AB, Hall A. Rho
GTPases: biochemistry and biology[J/OL]. Annu Rev Cell Dev Biol. 2005;21:247–69. Article CAS Google Scholar * Hodge RG, Ridley AJ. Regulating Rho GTPases and their regulators[J/OL]. Nat
Rev Mol Cell Biol. 2016;17:496–510. Article CAS Google Scholar * Hall A. Rho GTPases and the actin cytoskeleton[J/OL]. Science. 1998;279:509–14. Article CAS Google Scholar * De Lange
KM, Moutsianas L, Lee JC, Lamb CA, Luo Y, Kennedy NA, et al. Genome-wide association study implicates immune activation of multiple integrin genes in inflammatory bowel disease[J/OL]. Nat
Genet. 2017;49:256–61. Article Google Scholar * Fattouh R, Guo CH, Lam GY, Gareau MG, Ngan BY, Glogauer M, et al. Rac2-deficiency leads to exacerbated and protracted colitis in response to
Citrobacter rodentium infection[J/OL]. PloS One. 2013;8:e61629. Article CAS Google Scholar * Muise AM, Xu W, Guo CH, Walters TD, Wolters VM, Fattouh R, et al. NADPH oxidase complex and
IBD candidate gene studies: identification of a rare variant in NCF2 that results in reduced binding to RAC2[J/OL]. Gut. 2012;61:1028–35. Article CAS Google Scholar * Sironi M, Guerini
FR, Agliardi C, Biasin M, Cagliani R, Fumagalli M, et al. An evolutionary analysis of RAC2 identifies haplotypes associated with human autoimmune diseases[J/OL]. Mol Biol Evol.
2011;28:3319–29. Article CAS Google Scholar * Muise AM, Walters T, Xu W, Shen-Tu G, Guo CH, Fattouh R, et al. Single nucleotide polymorphisms that increase expression of the guanosine
triphosphatase RAC1 are associated with ulcerative colitis[J/OL]. Gastroenterology. 2011;141:633–41. Article CAS Google Scholar * Koifman E, Karban A, Mazor Y, Chermesh I, Waterman M,
Almog R, et al. Thiopurine effectiveness in patients with Crohn’s disease: a study of genetic and clinical predictive factors[J/OL]. Inflamm Bowel Dis. 2013;19:1639–44. Article Google
Scholar * Taher M, Ebrahimi Daryani N, Hedayat M, Eslamian M, Farhadi E, Mahmoudi M, et al. Association analysis of RAC1 single nucleotide polymorphisms with ulcerative colitis[J/OL]. Clin
Res Hepatol Gastroenterol. 2017;41:487–9. Article CAS Google Scholar * López-Posadas R, Becker C, Günther C, Tenzer S, Amann K, Billmeier U, et al. Rho-A prenylation and signaling link
epithelial homeostasis to intestinal inflammation[J/OL]. J Clin Investig. 2016;126:611–26. Article Google Scholar * Van Bodegraven AA, Curley CR, Hunt KA, Monsuur AJ, Linskens RK, Onnie
CM, et al. Genetic variation in myosin IXB is associated with ulcerative colitis[J/OL]. Gastroenterology. 2006;131:1768–74. Article Google Scholar * Latiano A, Palmieri O, Valvano MR,
D’incà R, Caprilli R, Cucchiara S, et al. The association of MYO9B gene in Italian patients with inflammatory bowel diseases[J/OL]. Aliment Pharmacol Ther. 2008;27:241–8. Article CAS
Google Scholar * Chandhoke SK, Mooseker MS. A role for myosin IXb, a motor-RhoGAP chimera, in epithelial wound healing and tight junction regulation[J/OL]. Mol Biol Cell. 2012;23:2468–80.
Article CAS Google Scholar * Zhang FL, Casey PJ. Influence of metal ions on substrate binding and catalytic activity of mammalian protein geranylgeranyltransferase type-I[J/OL]. Biochem
J. 1996;320:925–32. Article CAS Google Scholar * López-Posadas R, Fastancz P, Martínez-Sánchez LDC, Panteleev-Ivlev J, Thonn V, Kisseleva T, et al. Inhibiting PGGT1B Disrupts Function of
RHOA, Resulting in T-cell Expression of Integrin α4β7 and Development of Colitis in Mice[J/OL]. Gastroenterology. 2019;157:1293–309. Article Google Scholar * Wildenberg ME, Koelink PJ,
Diederen K, Te Velde AA, Wolfkamp SCS, Nuij VJ, et al. The ATG16L1 risk allele associated with Crohn’s disease results in a Rac1-dependent defect in dendritic cell migration that is
corrected by thiopurines[J/OL]. Mucosal. Immunology. 2017;10:352–60. CAS Google Scholar * Prins MMC, Giugliano FP, Van Roest M, Van De Graaf SFJ, Koelink PJ, Wildenberg ME. Thiopurines
correct the effects of autophagy impairment on intestinal healing - a potential role for ARHGAP18/RhoA[J/OL]. Dis Model Mech. 2021;14:dmm047233. Article CAS Google Scholar * Lemoine R,
Pachlopnik-Schmid J, Farin HF, Bigorgne A, Debré M, Sepulveda F, et al. Immune deficiency-related enteropathy-lymphocytopenia-alopecia syndrome results from tetratricopeptide repeat domain
7A deficiency[J/OL]. The. J Allergy Clin Immunol. 2014;134:1354–64. e6. Article CAS Google Scholar * Connelly TM, Sehgal R, Berg AS, Hegarty JP, Deiling S, Stewart DB, et al. Mutation in
TAGAP is protective of anal sepsis in ileocolic Crohn’s disease[J/OL]. Dis Colon Rectum. 2012;55:1145–52. Article Google Scholar * Lee M, Chang EB. Inflammatory Bowel Diseases (IBD) and
the Microbiome-Searching the Crime Scene for Clues[J/OL]. Gastroenterology. 2021;160:524–37. Article CAS Google Scholar * Maloy KJ, Powrie F. Intestinal homeostasis and its breakdown in
inflammatory bowel disease[J/OL]. Nature. 2011;474:298–306. Article CAS Google Scholar * Ladinsky MS, Araujo LP, Zhang X, Veltri J, Galan-Diez M, Soualhi S, et al. Endocytosis of
commensal antigens by intestinal epithelial cells regulates mucosal T cell homeostasis[J/OL]. Science. 2019;363:eaat4042. Article CAS Google Scholar * Nishino K, Nishida A, Inoue R,
Kawada Y, Ohno M, Sakai S, et al. Analysis of endoscopic brush samples identified mucosa-associated dysbiosis in inflammatory bowel disease[J/OL]. J Gastroenterol. 2018;53:95–106. Article
Google Scholar * Sokol H, Leducq V, Aschard H, Pham HP, Jegou S, Landman C, et al. Fungal microbiota dysbiosis in IBD[J/OL]. Gut. 2017;66:1039–48. Article CAS Google Scholar * Sartor RB,
Wu GD. Roles for Intestinal Bacteria, Viruses, and Fungi in Pathogenesis of Inflammatory Bowel Diseases and Therapeutic Approaches[J/OL]. Gastroenterology. 2017;152:327–39. e4. Article CAS
Google Scholar * Palmela C, Chevarin C, Xu Z, Torres J, Sevrin G, Hirten R, et al. Adherent-invasive Escherichia coli in inflammatory bowel disease[J/OL]. Gut. 2018;67:574–87. Article
CAS Google Scholar * Boyer L, Magoc L, Dejardin S, Cappillino M, Paquette N, Hinault C, et al. Pathogen-derived effectors trigger protective immunity via activation of the Rac2 enzyme and
the IMD or Rip kinase signaling pathway[J/OL]. Immunity. 2011;35:536–49. Article CAS Google Scholar * Wong ARC, Clements A, Raymond B, Crepin VF, Frankel G. The interplay between the
Escherichia coli Rho guanine nucleotide exchange factor effectors and the mammalian RhoGEF inhibitor EspH[J/OL]. mBio. 2012;3:e00250–11. Article CAS Google Scholar * Macdonald TT, Frankel
G, Dougan G, Goncalves NS, Simmons C. Host defences to Citrobacter rodentium[J/OL]. Int J Med Microbiol. 2003;293:87–93. Article CAS Google Scholar * Hardt WD, Chen LM, Schuebel KE,
Bustelo XR, Galán JES. typhimurium encodes an activator of Rho GTPases that induces membrane ruffling and nuclear responses in host cells[J/OL]. Cell. 1998;93:815–26. Article CAS Google
Scholar * Keestra AM, Winter MG, Auburger JJ, Frässle SP, Xavier MN, Winter SE, et al. Manipulation of small Rho GTPases is a pathogen-induced process detected by NOD1[J/OL]. Nature.
2013;496:233–7. Article CAS Google Scholar * Rodríguez C, Romero E, Garrido-Sanchez L, Alcaín-Martínez G, Andrade RJ, Taminiau B, et al. MICROBIOTA INSIGHTS IN CLOSTRIDIUM DIFFICILE
INFECTION AND INFLAMMATORY BOWEL DISEASE[J/OL]. Gut Microbes. 2020;12:1725220. Article Google Scholar * Berkes J, Viswanathan VK, Savkovic SD, Hecht G. Intestinal epithelial responses to
enteric pathogens: effects on the tight junction barrier, ion transport, and inflammation[J/OL]. Gut. 2003;52:439–51. Article CAS Google Scholar * Guan Q. A Comprehensive Review and
Update on the Pathogenesis of Inflammatory Bowel Disease[J/OL]. J Immunol Res. 2019;2019:7247238. Article Google Scholar * Nyström EEL, Martinez-Abad B, Arike L, Birchenough GMH, Nonnecke
EB, Castillo PA, et al. An intercrypt subpopulation of goblet cells is essential for colonic mucus barrier function[J/OL]. Science. 2021;372:eabb1590. Article Google Scholar * Wehkamp J,
Harder J, Weichenthal M, Mueller O, Herrlinger KR, Fellermann K, et al. Inducible and constitutive beta-defensins are differentially expressed in Crohn’s disease and ulcerative
colitis[J/OL]. Inflamm Bowel Dis. 2003;9:215–23. Article Google Scholar * Wehkamp J, Salzman NH, Porter E, Nuding S, Weichenthal M, Petras RE, et al. Reduced Paneth cell alpha-defensins in
ileal Crohn’s disease[J/OL]. Proc Natl Acad Sci USA. 2005;102:18129–34. Article CAS Google Scholar * Hopkins AM, Walsh SV, Verkade P, Boquet P, Nusrat A. Constitutive activation of Rho
proteins by CNF-1 influences tight junction structure and epithelial barrier function[J/OL]. J Cell Sci. 2003;116:725–42. Article CAS Google Scholar * Citalán-Madrid AF, Vargas-Robles H,
García-Ponce A, Shibayama M, Betanzos A, Nava P, et al. Cortactin deficiency causes increased RhoA/ROCK1-dependent actomyosin contractility, intestinal epithelial barrier dysfunction, and
disproportionately severe DSS-induced colitis[J/OL]. Mucosal. Immunology. 2017;10:1237–47. Google Scholar * Yang Y, Ma Y, Shi C, Chen H, Zhang H, Chen N, et al. Overexpression of miR-21 in
patients with ulcerative colitis impairs intestinal epithelial barrier function through targeting the Rho GTPase RhoB[J/OL]. Biochem Biophys Res Commun. 2013;434:746–52. Article CAS Google
Scholar * Shi C, Liang Y, Yang J, Xia Y, Chen H, Han H, et al. MicroRNA-21 knockout improve the survival rate in DSS induced fatal colitis through protecting against inflammation and
tissue injury[J/OL]. PloS One. 2013;8:e66814. Article CAS Google Scholar * Bruewer M, Luegering A, Kucharzik T, Parkos CA, Madara JL, Hopkins AM, et al. Proinflammatory cytokines disrupt
epithelial barrier function by apoptosis-independent mechanisms[J/OL]. J Immunol. 2003;171:6164–72. Article CAS Google Scholar * Utech M, Ivanov AI, Samarin SN, Bruewer M, Turner JR,
Mrsny RJ, et al. Mechanism of IFN-gamma-induced endocytosis of tight junction proteins: myosin II-dependent vacuolarization of the apical plasma membrane[J/OL]. Mol Biol Cell.
2005;16:5040–52. Article CAS Google Scholar * Jin S, Ray RM, Johnson LR. Rac1 mediates intestinal epithelial cell apoptosis via JNK[J/OL]. American Journal of Physiology. Gastrointest
Liver Physiol. 2006;291:G1137–47. Article CAS Google Scholar * Brito GAC, Fujji J, Carneiro-Filho BA, Lima AAM, Obrig T, Guerrant RL. Mechanism of Clostridium difficile toxin A-induced
apoptosis in T84 cells[J/OL]. The. J Infect Dis. 2002;186:1438–47. Article CAS Google Scholar * Fiorentini C, Fabbri A, Falzano L, Fattorossi A, Matarrese P, Rivabene R, et al.
Clostridium difficile toxin B induces apoptosis in intestinal cultured cells[J/OL]. Infect Immun. 1998;66:2660–5. Article CAS Google Scholar * Zheng XB, Liu HS, Zhang LJ, Liu XH, Zhong
XL, Zhou C, et al. Engulfment and Cell Motility Protein 1 Protects Against DSS-induced Colonic Injury in Mice via Rac1 Activation[J/OL]. J Crohn’s Colitis. 2019;13:100–14. Article Google
Scholar * Liu L, Zhuang R, Xiao L, Chung HK, Luo J, Turner DJ, et al. HuR Enhances Early Restitution of the Intestinal Epithelium by Increasing Cdc42 Translation[J/OL]. Mol Cell Biol.
2017;37:e00574–16. Article CAS Google Scholar * Giesemann T, Guttenberg G, Aktories K. Human alpha-defensins inhibit Clostridium difficile toxin B[J/OL]. Gastroenterology.
2008;134:2049–58. Article CAS Google Scholar * Fischer S, Ückert AK, Landenberger M, Papatheodorou P, Hoffmann-Richter C, Mittler AK, et al. Human peptide α-defensin-1 interferes with
Clostridioides difficile toxins TcdA, TcdB, and CDT[J/OL]. FASEB J. 2020;34:6244–61. Article CAS Google Scholar * Sheng Q, Lv Z, Cai W, Song H, Qian L, Mu H, et al. Human β-defensin-3
promotes intestinal epithelial cell migration and reduces the development of necrotizing enterocolitis in a neonatal rat model[J/OL]. Pediatr Res. 2014;76:269–79. Article CAS Google
Scholar * Coombes JL, Powrie F. Dendritic cells in intestinal immune regulation[J/OL]. Nat Rev Immunol. 2008;8:435–46. Article CAS Google Scholar * Na YR, Stakenborg M, Seok SH, Matteoli
G. Macrophages in intestinal inflammation and resolution: a potential therapeutic target in IBD[J/OL]. Nature Reviews. Gastroenterol Hepatol. 2019;16:531–43. CAS Google Scholar * Baumgart
DC, Carding SR. Inflammatory bowel disease: cause and immunobiology[J/OL]. Lancet. 2007;369:1627–40. Article CAS Google Scholar * Steinbach EC, Plevy SE. The role of macrophages and
dendritic cells in the initiation of inflammation in IBD[J/OL]. Inflamm Bowel Dis. 2014;20:166–75. Article Google Scholar * Garrett WS, Chen LM, Kroschewski R, Ebersold M, Turley S,
Trombetta S, et al. Developmental control of endocytosis in dendritic cells by Cdc42[J/OL]. Cell. 2000;102:325–34. Article CAS Google Scholar * West MA, Prescott AR, Eskelinen EL, Ridley
AJ, Watts C. Rac is required for constitutive macropinocytosis by dendritic cells but does not control its downregulation[J/OL]. Curr Biol. 2000;10:839–48. Article CAS Google Scholar *
Zhang J, Guo J, Dzhagalov I, He YW. An essential function for the calcium-promoted Ras inactivator in Fcgamma receptor-mediated phagocytosis[J/OL]. Nat Immunol. 2005;6:911–9. Article CAS
Google Scholar * Niedergang F, Chavrier P. Regulation of phagocytosis by Rho GTPases[J/OL]. Curr Top Microbiol Immunol. 2005;291:43–60. CAS Google Scholar * Mao Y, Finnemann SC.
Regulation of phagocytosis by Rho GTPases[J/OL]. Small GTPases. 2015;6:89–99. Article CAS Google Scholar * Egami Y, Kawai K, Araki N. RhoC regulates the actin remodeling required for
phagosome formation during FcγR-mediated phagocytosis[J/OL]. J Cell Sci. 2017;130:4168–79. CAS Google Scholar * Geng J, Shi Y, Zhang J, Yang B, Wang P, Yuan W, et al. TLR4 signalling via
Piezo1 engages and enhances the macrophage mediated host response during bacterial infection[J/OL]. Nature. Communications. 2021;12:3519. CAS Google Scholar * Tzircotis G, Braga VMM, Caron
E. RhoG is required for both FcγR- and CR3-mediated phagocytosis[J/OL]. J Cell Sci. 2011;124:2897–902. Article CAS Google Scholar * Leverrier Y, Ridley AJ. Requirement for Rho GTPases
and PI 3-kinases during apoptotic cell phagocytosis by macrophages[J/OL]. Curr Biol. 2001;11:195–9. Article CAS Google Scholar * Nakaya M, Tanaka M, Okabe Y, Hanayama R, Nagata S.
Opposite effects of rho family GTPases on engulfment of apoptotic cells by macrophages[J/OL]. J Biol Chem. 2006;281:8836–42. Article CAS Google Scholar * Burns S, Thrasher AJ, Blundell
MP, Machesky L, Jones GE. Configuration of human dendritic cell cytoskeleton by Rho GTPases, the WAS protein, and differentiation[J/OL]. Blood. 2001;98:1142–9. Article CAS Google Scholar
* Swetman CA, Leverrier Y, Garg R, Gan CHV, Ridley AJ, Katz DR, et al. Extension, retraction and contraction in the formation of a dendritic cell dendrite: distinct roles for Rho
GTPases[J/OL]. Eur J Immunol. 2002;32:2074–83. Article CAS Google Scholar * Erwig LP, Mcphilips KA, Wynes MW, Ivetic A, Ridley AJ, Henson PM. Differential regulation of phagosome
maturation in macrophages and dendritic cells mediated by Rho GTPases and ezrin-radixin-moesin (ERM) proteins[J/OL]. Proc Natl Acad Sci USA. 2006;103:12825–30. Article CAS Google Scholar
* Wu W, Hsu YMS, Bi L, Songyang Z, Lin X. CARD9 facilitates microbe-elicited production of reactive oxygen species by regulating the LyGDI-Rac1 complex[J/OL]. Nat Immunol. 2009;10:1208–14.
Article CAS Google Scholar * Li Y, Kim JG, Kim HJ, Moon MY, Lee JY, Kim J, et al. Small GTPases Rap1 and RhoA regulate superoxide formation by Rac1 GTPases activation during the
phagocytosis of IgG-opsonized zymosans in macrophages[J/OL]. Free Radic Biol Med. 2012;52:1796–805. Article CAS Google Scholar * Ridley AJ. Rho proteins, PI 3-kinases, and
monocyte/macrophage motility[J/OL]. FEBS Lett. 2001;498:168–71. Article CAS Google Scholar * Infante E, Ridley AJ. Roles of Rho GTPases in leucocyte and leukaemia cell transendothelial
migration[J/OL]. Philos Trans R Soc Lond Ser B, Biol Sci. 2013;368:20130013. Article Google Scholar * Levine AP, Segal AW. What is wrong with granulocytes in inflammatory bowel
diseases?[J/OL]. Dig Dis. 2013;31:321–7. Article Google Scholar * Wéra O, Lancellotti P, Oury C. The Dual Role of Neutrophils in Inflammatory Bowel Diseases[J/OL]. J Clin Med. 2016;5:118.
Article Google Scholar * Petri B, Sanz MJ. Neutrophil chemotaxis[J/OL]. Cell Tissue Res. 2018;371:425–36. Article CAS Google Scholar * Werner E. GTPases and reactive oxygen species:
switches for killing and signaling[J/OL]. J Cell Sci. 2004;117:143–53. Article CAS Google Scholar * Lacy P. The role of Rho GTPases and SNAREs in mediator release from granulocytes[J/OL].
Pharmacol Ther. 2005;107:358–76. Article CAS Google Scholar * Tybulewicz VLJ, Henderson RB. Rho family GTPases and their regulators in lymphocytes[J/OL]. Nat Rev Immunol. 2009;9:630–44.
Article CAS Google Scholar * Cernuda-Morollón E, Millán J, Shipman M, Marelli-Berg FM, Ridley AJ. Rac activation by the T-cell receptor inhibits T cell migration[J/OL]. PloS One.
2010;5:e12393. Article Google Scholar * García-Bernal D, Wright N, Sotillo-Mallo E, Nombela-Arrieta C, Stein JV, Bustelo XR, et al. Vav1 and Rac control chemokine-promoted T lymphocyte
adhesion mediated by the integrin alpha4beta1[J/OL]. Mol Biol Cell. 2005;16:3223–35. Article Google Scholar * Vielkind S, Gallagher-Gambarelli M, Gomez M, Hinton HJ, Cantrell DA. Integrin
regulation by RhoA in thymocytes[J/OL]. J Immunol. 2005;175:350–7. Article CAS Google Scholar * Faroudi M, Hons M, Zachacz A, Dumont C, Lyck R, Stein JV, et al. Critical roles for Rac
GTPases in T-cell migration to and within lymph nodes[J/OL]. Blood. 2010;116:5536–47. Article CAS Google Scholar * Baker CM, Comrie WA, Hyun YM, Chung HL, Fedorchuk CA, Lim K, et al.
Opposing roles for RhoH GTPase during T-cell migration and activation[J/OL]. Proc Natl Acad Sci USA. 2012;109:10474–9. Article CAS Google Scholar * Singleton KL, Roybal KT, Sun Y, Fu G,
Gascoigne NRJ, Van Oers NSC, et al. Spatiotemporal patterning during T cell activation is highly diverse[J/OL]. Sci Signal. 2009;2:ra15. Article Google Scholar * Faure S, Salazar-Fontana
LI, Semichon M, Tybulewicz VLJ, Bismuth G, Trautmann A, et al. ERM proteins regulate cytoskeleton relaxation promoting T cell-APC conjugation[J/OL]. Nat Immunol. 2004;5:272–9. Article CAS
Google Scholar * Delaguillaumie A, Lagaudrière-Gesbert C, Popoff MR, Conjeaud H. Rho GTPases link cytoskeletal rearrangements and activation processes induced via the tetraspanin CD82 in T
lymphocytes[J/OL]. J Cell Sci. 2002;115:433–43. Article CAS Google Scholar * Salazar-Fontana LI, Barr V, Samelson LE, Bierer BE. CD28 engagement promotes actin polymerization through the
activation of the small Rho GTPase Cdc42 in human T cells[J/OL]. J Immunol. 2003;171:2225–32. Article CAS Google Scholar * Jacinto E, Werlen G, Karin M. Cooperation between Syk and Rac1
leads to synergistic JNK activation in T lymphocytes[J/OL]. Immunity. 1998;8:31–41. Article CAS Google Scholar * Marinari B, Costanzo A, Viola A, Michel F, Mangino G, Acuto O, et al. Vav
cooperates with CD28 to induce NF-kappaB activation via a pathway involving Rac-1 and mitogen-activated kinase kinase 1[J/OL]. Eur J Immunol. 2002;32:447–56. Article CAS Google Scholar *
Arrieumerlou C, Randriamampita C, Bismuth G, Trautmann A. Rac is involved in early TCR signaling[J/OL]. J Immunol. 2000;165:3182–9. Article CAS Google Scholar * Oh-Hora M, Rao A. Calcium
signaling in lymphocytes[J/OL]. Curr Opin Immunol. 2008;20:250–8. Article CAS Google Scholar * Acuto O, Cantrell D. T cell activation and the cytoskeleton[J/OL]. Annu Rev Immunol.
2000;18:165–84. Article CAS Google Scholar * Angkachatchai V, Finkel TH. ADP-ribosylation of rho by C3 ribosyltransferase inhibits IL-2 production and sustained calcium influx in
activated T cells[J]. J Immunol. 1999;163:3819–25. Article CAS Google Scholar * Yang JQ, Kalim KW, Li Y, Zhang S, Hinge A, Filippi MD, et al. RhoA orchestrates glycolysis for TH2 cell
differentiation and allergic airway inflammation[J/OL]. J Allergy Clin Immunol. 2016;137:231–45. e4. Article CAS Google Scholar * Guo F, Hildeman D, Tripathi P, Velu CS, Grimes HL, Zheng
Y. Coordination of IL-7 receptor and T-cell receptor signaling by cell-division cycle 42 in T-cell homeostasis[J/OL]. Proc Natl Acad Sci USA. 2010;107:18505–10. Article CAS Google Scholar
* Ahmad Mokhtar AM, Salikin NH, Haron AS, Amin-Nordin S, Hashim IF, Mohd Zaini Makhtar M, et al. RhoG’s Role in T Cell Activation and Function[J/OL]. Front Immunol. 2022;13:845064. Article
Google Scholar * Kalim KW, Yang JQ, Li Y, Meng Y, Zheng Y, Guo F. Reciprocal Regulation of Glycolysis-Driven Th17 Pathogenicity and Regulatory T Cell Stability by Cdc42[J/OL]. J Immunol.
2018;200:2313–26. Article CAS Google Scholar * Yang JQ, Kalim KW, Li Y, Duan X, Nguyen P, Khurana Hershey GK, et al. Rational targeting Cdc42 restrains Th2 cell differentiation and
prevents allergic airway inflammation[J/OL]. Clin Exp Allergy. 2019;49:92–107. Article CAS Google Scholar * Yang JQ, Kalim KW, Li Y, Zheng Y, Guo F. Ablation of RhoA impairs Th17 cell
differentiation and alleviates house dust mite-triggered allergic airway inflammation[J/OL]. J Leukoc Biol. 2019;106:1139–51. Article CAS Google Scholar * Dunn SE, Youssef S, Goldstein
MJ, Prod’homme T, Weber MS, Zamvil SS, et al. Isoprenoids determine Th1/Th2 fate in pathogenic T cells, providing a mechanism of modulation of autoimmunity by atorvastatin[J/OL]. J Exp Med.
2006;203:401–12. Article CAS Google Scholar * Li B, Yu H, Zheng W, Voll R, Na S, Roberts AW, et al. Role of the guanosine triphosphatase Rac2 in T helper 1 cell differentiation[J/OL].
Science. 2000;288:2219–22. Article CAS Google Scholar * Li X, Bu X, Lu B, Avraham H, Flavell RA, Lim B. The hematopoiesis-specific GTP-binding protein RhoH is GTPase deficient and
modulates activities of other Rho GTPases by an inhibitory function[J/OL]. Mol Cell Biol. 2002;22:1158–71. Article CAS Google Scholar * Tang HC, Lai YY, Zheng J, Jiang HY, Xu G.
miR-223-3p Inhibits Antigen Endocytosis and Presentation and Promotes the Tolerogenic Potential of Dendritic Cells through Targeting Mannose Receptor Signaling and Rhob[J/OL]. J Immunol Res.
2020;2020:1379458. Article Google Scholar * Na S, Li B, Grewal IS, Enslen H, Davis RJ, Hanke JH, et al. Expression of activated CDC42 induces T cell apoptosis in thymus and peripheral
lymph organs via different pathways[J/OL]. Oncogene. 1999;18:7966–74. Article CAS Google Scholar * Subauste MC, Von Herrath M, Benard V, Chamberlain CE, Chuang TH, Chu K, et al. Rho
family proteins modulate rapid apoptosis induced by cytotoxic T lymphocytes and Fas[J/OL]. J Biol Chem. 2000;275:9725–33. Article CAS Google Scholar * Hébert M, Potin S, Sebbagh M,
Bertoglio J, Bréard J, Hamelin J. Rho-ROCK-dependent ezrin-radixin-moesin phosphorylation regulates Fas-mediated apoptosis in Jurkat cells[J/OL]. J Immunol. 2008;181:5963–73. Article Google
Scholar * Brinkkoetter PT, Gottmann U, Schulte J, Van Der Woude FJ, Braun C, Yard BA. Atorvastatin interferes with activation of human CD4(+) T cells via inhibition of small guanosine
triphosphatase (GTPase) activity and caspase-independent apoptosis[J/OL]. Clin Exp Immunol. 2006;146:524–32. Article CAS Google Scholar * Tiede I, Fritz G, Strand S, Poppe D, Dvorsky R,
Strand D, et al. CD28-dependent Rac1 activation is the molecular target of azathioprine in primary human CD4+ T lymphocytes[J/OL]. J Clin Investig. 2003;111:1133–45. Article CAS Google
Scholar * Poppe D, Tiede I, Fritz G, Becker C, Bartsch B, Wirtz S, et al. Azathioprine suppresses ezrin-radixin-moesin-dependent T cell-APC conjugation through inhibition of Vav guanosine
exchange activity on Rac proteins[J/OL]. J Immunol. 2006;176:640–51. Article CAS Google Scholar * Neurath MF. Cytokines in inflammatory bowel disease[J/OL]. Nat Rev Immunol.
2014;14:329–42. Article CAS Google Scholar * Dong LM, Chen XW, He XX, Jiang XP, Wu F. Cell division cycle protein 42 regulates the inflammatory response in mice bearing inflammatory bowel
disease[J/OL]. Artif Cells, Nanomed, Biotechnol. 2019;47:1833–8. Article CAS Google Scholar * Tang WJ, Peng KY, Tang ZF, Wang YH, Xue AJ, Huang Y. MicroRNA-15a - cell division cycle 42
signaling pathway in pathogenesis of pediatric inflammatory bowel disease[J/OL]. World J Gastroenterol. 2018;24:5234–45. Article CAS Google Scholar * Segain JP, Raingeard DE, La Blétière
D, Sauzeau V, Bourreille A, Hilaret G, et al. Rho kinase blockade prevents inflammation via nuclear factor kappa B inhibition: evidence in Crohn’s disease and experimental colitis[J/OL].
Gastroenterology. 2003;124:1180–7. Article CAS Google Scholar * Johnson LA, Rodansky ES, Haak AJ, Larsen SD, Neubig RR, Higgins PDR. Novel Rho/MRTF/SRF inhibitors block matrix-stiffness
and TGF-β-induced fibrogenesis in human colonic myofibroblasts[J/OL]. Inflamm Bowel Dis. 2014;20:154–65. Article Google Scholar * Horowitz S, Binion DG, Nelson VM, Kanaa Y, Javadi P,
Lazarova Z, et al. Increased arginase activity and endothelial dysfunction in human inflammatory bowel disease[J/OL]. Am J Physiol Gastrointest Liver Physiol. 2007;292:G1323–1336. Article
CAS Google Scholar * Seidelin JB, Larsen S, Linnemann D, Vainer B, Coskun M, Troelsen JT, et al. Cellular inhibitor of apoptosis protein 2 controls human colonic epithelial restitution,
migration, and Rac1 activation[J/OL]. Am J Physiol Gastrointest Liver Physiol. 2015;308:G92–99. Article CAS Google Scholar * Parikh K, Zhou L, Somasundaram R, Fuhler GM, Deuring JJ,
Blokzijl T, et al. Suppression of p21Rac signaling and increased innate immunity mediate remission in Crohn’s disease[J/OL]. Sci Transl Med. 2014;6:233ra53. Article Google Scholar *
Marinković G, Hamers AAJ, De Vries CJM, De Waard V. 6-Mercaptopurine reduces macrophage activation and gut epithelium proliferation through inhibition of GTPase Rac1[J/OL]. Inflamm Bowel
Dis. 2014;20:1487–95. Article Google Scholar * Liu G, Gu K, Wang F, Jia G, Zhao H, Chen X, et al. Tryptophan Ameliorates Barrier Integrity and Alleviates the Inflammatory Response to
Enterotoxigenic Escherichia coli K88 Through the CaSR/Rac1/PLC-γ1 Signaling Pathway in Porcine Intestinal Epithelial Cells[J/OL]. Front Immunol. 2021;12:748497. Article CAS Google Scholar
* Piovani D, Danese S, Peyrin-Biroulet L, Nikolopoulos GK, Lytras T, Bonovas S. Environmental Risk Factors for Inflammatory Bowel Diseases: An Umbrella Review of Meta-analyses[J/OL].
Gastroenterology. 2019;157:647–59. e4. Article Google Scholar * Kanauchi Y, Yamamoto T, Yoshida M, Zhang Y, Lee J, Hayashi S, et al. Cholinergic anti-inflammatory pathway ameliorates
murine experimental Th2-type colitis by suppressing the migration of plasmacytoid dendritic cells[J/OL]. Sci Rep. 2022;12:54. Article CAS Google Scholar * Penna G, Fibbi B, Amuchastegui
S, Corsiero E, Laverny G, Silvestrini E, et al. The vitamin D receptor agonist elocalcitol inhibits IL-8-dependent benign prostatic hyperplasia stromal cell proliferation and inflammatory
response by targeting the RhoA/Rho kinase and NF-kappaB pathways[J/OL]. Prostate. 2009;69:480–93. Article CAS Google Scholar * Qian X, Zhu M, Qian W, Song J. Vitamin D attenuates
myocardial ischemia-reperfusion injury by inhibiting inflammation via suppressing the RhoA/ROCK/NF-ĸB pathway[J/OL]. Biotechnol Appl Biochem. 2019;66:850–7. Article CAS Google Scholar *
Aslan A, Triadafilopoulos G. Fish oil fatty acid supplementation in active ulcerative colitis: a double-blind, placebo-controlled, crossover study[J]. Am J Gastroenterol. 1992;87:432–7. CAS
Google Scholar * Hou JK, Abraham B, El-Serag H. Dietary intake and risk of developing inflammatory bowel disease: a systematic review of the literature[J/OL]. Am J Gastroenterol.
2011;106:563–73. Article CAS Google Scholar * Takeuchi O, Akira S. Pattern recognition receptors and inflammation[J/OL]. Cell. 2010;140:805–20. Article CAS Google Scholar * Kordjazy N,
Haj-Mirzaian A, Haj-Mirzaian A, Rohani MM, Gelfand EW, Rezaei N, et al. Role of toll-like receptors in inflammatory bowel disease[J/OL]. Pharmacol Res. 2018;129:204–15. Article CAS Google
Scholar * Arbibe L, Mira JP, Teusch N, Kline L, Guha M, Mackman N, et al. Toll-like receptor 2-mediated NF-kappa B activation requires a Rac1-dependent pathway[J/OL]. Nat Immunol.
2000;1:533–40. Article CAS Google Scholar * Teusch N, Lombardo E, Eddleston J, Knaus UG. The low molecular weight GTPase RhoA and atypical protein kinase Czeta are required for
TLR2-mediated gene transcription[J/OL]. J Immunol. 2004;173:507–14. Article CAS Google Scholar * Schmeck B, Huber S, Moog K, Zahlten J, Hocke AC, Opitz B, et al. Pneumococci induced TLR-
and Rac1-dependent NF-kappaB-recruitment to the IL-8 promoter in lung epithelial cells[J/OL]. Am J Physiol Lung Cell Mol Physiol. 2006;290:L730–L737. Article CAS Google Scholar * Choi SH,
Harkewicz R, Lee JH, Boullier A, Almazan F, Li AC, et al. Lipoprotein accumulation in macrophages via toll-like receptor-4-dependent fluid phase uptake[J/OL]. Circ Res. 2009;104:1355–63.
Article CAS Google Scholar * Kong L, Ge BX. MyD88-independent activation of a novel actin-Cdc42/Rac pathway is required for Toll-like receptor-stimulated phagocytosis[J/OL]. Cell Res.
2008;18:745–55. Article CAS Google Scholar * Gotoh K, Tanaka Y, Nishikimi A, Nakamura R, Yamada H, Maeda N, et al. Selective control of type I IFN induction by the Rac activator DOCK2
during TLR-mediated plasmacytoid dendritic cell activation[J/OL]. J Exp Med. 2010;207:721–30. Article CAS Google Scholar * Wang Z, Fayngerts S, Wang P, Sun H, Johnson DS, Ruan Q, et al.
TIPE2 protein serves as a negative regulator of phagocytosis and oxidative burst during infection[J/OL]. Proc Natl Acad Sci USA. 2012;109:15413–8. Article CAS Google Scholar * Liu S,
Huang L, Lin Z, Hu Y, Chen R, Wang L, et al. RhoB induces the production of proinflammatory cytokines in TLR-triggered macrophages[J/OL]. Mol Immunol. 2017;87:200–6. Article CAS Google
Scholar * Pulecio J, Petrovic J, Prete F, Chiaruttini G, Lennon-Dumenil AM, Desdouets C, et al. Cdc42-mediated MTOC polarization in dendritic cells controls targeted delivery of cytokines
at the immune synapse[J/OL]. J Exp Med. 2010;207:2719–32. Article CAS Google Scholar * Rubino SJ, Selvanantham T, Girardin SE, Philpott DJ. Nod-like receptors in the control of intestinal
inflammation[J/OL]. Curr Opin Immunol. 2012;24:398–404. Article CAS Google Scholar * Keestra AM, Bäumler AJ. Detection of enteric pathogens by the nodosome[J/OL]. Trends Immunol.
2014;35:123–30. Article CAS Google Scholar * Eitel J, Krüll M, Hocke AC, N’guessan PD, Zahlten J, Schmeck B, et al. Beta-PIX and Rac1 GTPase mediate trafficking and negative regulation of
NOD2[J/OL]. J Immunol. 2008;181:2664–71. Article CAS Google Scholar * Dufies O, Doye A, Courjon J, Torre C, Michel G, Loubatier C, et al. Escherichia coli Rho GTPase-activating toxin
CNF1 mediates NLRP3 inflammasome activation via p21-activated kinases-1/2 during bacteraemia in mice[J/OL]. Nat Microbiol. 2021;6:401–12. Article CAS Google Scholar * Xie S, Yang T, Wang
Z, Li M, Ding L, Hu X, et al. Astragaloside IV attenuates sepsis-induced intestinal barrier dysfunction via suppressing RhoA/NLRP3 inflammasome signaling[J/OL]. Int Immunopharmacol.
2020;78:106066. Article CAS Google Scholar * Ying C, Zhou Z, Dai J, Wang M, Xiang J, Sun D, et al. Activation of the NLRP3 inflammasome by RAC1 mediates a new mechanism in diabetic
nephropathy[J/OL]. Inflamm Res. 2022;71:191–204. Article CAS Google Scholar * Yan Y, Lu K, Ye T, Zhang Z. MicroRNA‑223 attenuates LPS‑induced inflammation in an acute lung injury model
via the NLRP3 inflammasome and TLR4/NF‑κB signaling pathway via RHOB[J/OL]. Int J Mol Med. 2019;43:1467–77. CAS Google Scholar * Rebane A, Akdis CA. MicroRNAs: Essential players in the
regulation of inflammation[J/OL]. J Allergy Clin Immunol. 2013;132:15–26. Article CAS Google Scholar * Wu F, Zikusoka M, Trindade A, Dassopoulos T, Harris ML, Bayless TM, et al. MicroRNAs
are differentially expressed in ulcerative colitis and alter expression of macrophage inflammatory peptide-2 alpha[J/OL]. Gastroenterology. 2008;135:1624–35. e24. Article CAS Google
Scholar * Wu F, Zhang S, Dassopoulos T, Harris ML, Bayless TM, Meltzer SJ, et al. Identification of microRNAs associated with ileal and colonic Crohn’s disease[J/OL]. Inflamm Bowel Dis.
2010;16:1729–38. Article Google Scholar * Pekow JR, Dougherty U, Mustafi R, Zhu H, Kocherginsky M, Rubin DT, et al. miR-143 and miR-145 are downregulated in ulcerative colitis: putative
regulators of inflammation and protooncogenes[J/OL]. Inflamm Bowel Dis. 2012;18:94–100. Article Google Scholar * Fang K, Law IKM, Padua D, Sideri A, Huang V, Kevil CG, et al.
MicroRNA-31-3p Is Involved in Substance P (SP)-Associated Inflammation in Human Colonic Epithelial Cells and Experimental Colitis[J/OL]. Am J Pathol. 2018;188:586–99. Article CAS Google
Scholar * Zhou WY, Cai ZR, Liu J, Wang DS, Ju HQ, Xu RH. Circular RNA: metabolism, functions and interactions with proteins[J/OL]. Mol Cancer. 2020;19:172. Article CAS Google Scholar *
Xiao L, Ma XX, Luo J, Chung HK, Kwon MS, Yu TX, et al. Circular RNA CircHIPK3 Promotes Homeostasis of the Intestinal Epithelium by Reducing MicroRNA 29b Function[J/OL]. Gastroenterology.
2021;161:1303–17. e3. Article CAS Google Scholar * Danese S, Sans M, De La Motte C, Graziani C, West G, Phillips MH, et al. Angiogenesis as a novel component of inflammatory bowel disease
pathogenesis[J/OL]. Gastroenterology. 2006;130:2060–73. Article CAS Google Scholar * Merajver SD, Usmani SZ. Multifaceted role of Rho proteins in angiogenesis[J/OL]. J Mammary Gland Biol
Neoplasia. 2005;10:291–8. Article Google Scholar * Sulpice E, Ding S, Muscatelli-Groux B, Bergé M, Han ZC, Plouet J, et al. Cross-talk between the VEGF-A and HGF signalling pathways in
endothelial cells[J/OL]. Biol Cell. 2009;101:525–39. Article CAS Google Scholar * Pang X, Zhang L, Lai L, Chen J, Wu Y, Yi Z, et al. 1’-Acetoxychavicol acetate suppresses
angiogenesis-mediated human prostate tumor growth by targeting VEGF-mediated Src-FAK-Rho GTPase-signaling pathway[J/OL]. Carcinogenesis. 2011;32:904–12. Article CAS Google Scholar * Lai
Y, Shen Y, Liu XH, Zhang Y, Zeng Y, Liu YF. Interleukin-8 induces the endothelial cell migration through the activation of phosphoinositide 3-kinase-Rac1/RhoA pathway[J/OL]. Int J Biol Sci.
2011;7:782–91. Article CAS Google Scholar * Turk HF, Monk JM, Fan YY, Callaway ES, Weeks B, Chapkin RS. Inhibitory effects of omega-3 fatty acids on injury-induced epidermal growth factor
receptor transactivation contribute to delayed wound healing[J/OL]. Am J Physiol Cell Physiol. 2013;304:C905–917. Article CAS Google Scholar * Maldonado MDM, Dharmawardhane S. Targeting
Rac and Cdc42 GTPases in Cancer[J/OL]. Cancer Res. 2018;78:3101–11. Article CAS Google Scholar * Budzyn K, Marley PD, Sobey CG. Targeting Rho and Rho-kinase in the treatment of
cardiovascular disease[J/OL]. Trends Pharmacol Sci. 2006;27:97–104. Article CAS Google Scholar * Aguilar BJ, Zhu Y, Lu Q. Rho GTPases as therapeutic targets in Alzheimer’s disease[J/OL].
Alzheimer’s Res Ther. 2017;9:97. Article Google Scholar * Martin TE, Powell CT, Wang Z, Bhattacharyya S, Walsh-Reitz MM, Agarwal K, et al. A novel mitogenic protein that is highly
expressed in cells of the gastric antrum mucosa[J/OL]. Am J Physiol Gastrointest Liver Physiol. 2003;285:G332–43. Article CAS Google Scholar * Chen P, Bakke D, Kolodziej L, Lodolce J,
Weber CR, Boone DL, et al. Antrum Mucosal Protein-18 Peptide Targets Tight Junctions to Protect and Heal Barrier Structure and Function in Models of Inflammatory Bowel Disease[J/OL]. Inflamm
Bowel Dis. 2015;21:2393–402. Google Scholar * Chen P, Kartha S, Bissonnette M, Hart J, Toback FG. AMP-18 facilitates assembly and stabilization of tight junctions to protect the colonic
mucosal barrier[J/OL]. Inflamm Bowel Dis. 2012;18:1749–59. Article Google Scholar * D’haens G, Van Deventer S, Van Hogezand R, Chalmers D, Kothe C, Baert F, et al. Endoscopic and
histological healing with infliximab anti-tumor necrosis factor antibodies in Crohn’s disease: A European multicenter trial[J/OL]. Gastroenterology. 1999;116:1029–34. Article Google Scholar
* Rutgeerts P, D’haens G, Targan S, Vasiliauskas E, Hanauer SB, Present DH, et al. Efficacy and safety of retreatment with anti-tumor necrosis factor antibody (infliximab) to maintain
remission in Crohn’s disease[J/OL]. Gastroenterology. 1999;117:761–9. Article CAS Google Scholar * Bouguen G, Roblin X, Bourreille A, Feier L, Filippi J, Nancey S, et al. Infliximab for
refractory ulcerative proctitis[J/OL]. Aliment Pharmacol Ther. 2010;31:1178–85. Article CAS Google Scholar * Caviglia R, Ribolsi M, Rizzi M, Emerenziani S, Annunziata ML, Cicala M.
Maintenance of remission with infliximab in inflammatory bowel disease: efficacy and safety long-term follow-up[J/OL]. World J Gastroenterol. 2007;13:5238–44. Article CAS Google Scholar *
Liu L, Liu Q, Chang J, Dong X, Ma W. Cell division control 42 elevates during infliximab therapy, and its increment relates to treatment response in ulcerative colitis patients[J/OL]. J
Clin Lab Anal. 2022;36:e24477. CAS Google Scholar * Gao Y, Dickerson JB, Guo F, Zheng J, Zheng Y. Rational design and characterization of a Rac GTPase-specific small molecule
inhibitor[J/OL]. Proc Natl Acad Sci USA. 2004;101:7618–23. Article CAS Google Scholar * Nielsen OH, Vainer B, Rask-Madsen J. Review article: the treatment of inflammatory bowel disease
with 6-mercaptopurine or azathioprine[J/OL]. Aliment Pharmacol Ther. 2001;15:1699–708. Article CAS Google Scholar * De Boer NKH, Peyrin-Biroulet L, Jharap B, Sanderson JD, Meijer B,
Atreya I, et al. Thiopurines in Inflammatory Bowel Disease: New Findings and Perspectives[J/OL]. J Crohn’s Colitis. 2018;12:610–20. Article Google Scholar * Seinen ML, Van Nieuw Amerongen
GP, De Boer NKH, Mulder CJJ, Van Bezu J, Van Bodegraven AA. Rac1 as a Potential Pharmacodynamic Biomarker for Thiopurine Therapy in Inflammatory Bowel Disease[J/OL]. Ther Drug Monit.
2016;38:621. Article CAS Google Scholar * Yusung S, Mcgovern D, Lin L, Hommes D, Lagishetty V, Braun J. NK cells are biologic and biochemical targets of 6-mercaptopurine in Crohn’s
disease patients[J/OL]. Clin Immunol. 2017;175:82–90. Article CAS Google Scholar * Wang CY, Liu PY, Liao JK. Pleiotropic effects of statin therapy: molecular mechanisms and clinical
results[J/OL]. Trends Mol Med. 2008;14:37–44. Article CAS Google Scholar * Grip O, Janciauskiene S, Bredberg A. Use of atorvastatin as an anti-inflammatory treatment in Crohn’s
disease[J/OL]. Br J Pharmacol. 2008;155:1085–92. Article CAS Google Scholar * Chardin P, Boquet P, Madaule P, Popoff MR, Rubin EJ, Gill DM. The mammalian G protein rhoC is ADP-ribosylated
by Clostridium botulinum exoenzyme C3 and affects actin microfilaments in Vero cells[J/OL]. EMBO J. 1989;8:1087–92. Article CAS Google Scholar * Uehata M, Ishizaki T, Satoh H, Ono T,
Kawahara T, Morishita T, et al. Calcium sensitization of smooth muscle mediated by a Rho-associated protein kinase in hypertension[J/OL]. Nature. 1997;389:990–4. Article CAS Google Scholar
* Guzman JR, Koo JS, Goldsmith JR, Mühlbauer M, Narula A, Jobin C. Oxymatrine prevents NF-κB nuclear translocation and ameliorates acute intestinal inflammation[J/OL]. Sci Rep.
2013;3:1629. Article Google Scholar * Choi YJ, Koo JB, Kim HY, Seo JW, Lee EJ, Kim WR, et al. Umbilical cord/placenta-derived mesenchymal stem cells inhibit fibrogenic activation in human
intestinal myofibroblasts via inhibition of myocardin-related transcription factor A[J/OL]. Stem Cell Res Ther. 2019;10:291. Article Google Scholar * Naito Y, Takagi T, Katada K, Uchiyama
K, Kuroda M, Kokura S, et al. Partially hydrolyzed guar gum down-regulates colonic inflammatory response in dextran sulfate sodium-induced colitis in mice[J/OL]. J Nutr Biochem.
2006;17:402–9. Article CAS Google Scholar * Wang Y, Shou Z, Fan H, Xu M, Chen Q, Tang Q, et al. Protective effects of oxymatrine against DSS-induced acute intestinal inflammation in mice
via blocking the RhoA/ROCK signaling pathway[J/OL]. Biosci Rep. 2019;39:BSR20182297. Article CAS Google Scholar * Horii Y, Uchiyama K, Toyokawa Y, Hotta Y, Tanaka M, Yasukawa Z, et al.
Partially hydrolyzed guar gum enhances colonic epithelial wound healing via activation of RhoA and ERK1/2[J/OL]. Food Funct. 2016;7:3176–83. Article CAS Google Scholar * Veeraperumal S,
Qiu HM, Tan CS, Ng ST, Zhang W, Tang S, et al. Restitution of epithelial cells during intestinal mucosal wound healing: The effect of a polysaccharide from the sclerotium of Lignosus
rhinocerotis (Cooke) Ryvarden[J/OL]. J Ethnopharmacol. 2021;274:114024. Article CAS Google Scholar * Bartok B, Hammaker D, Firestein GS. Phosphoinositide 3-kinase δ regulates migration
and invasion of synoviocytes in rheumatoid arthritis[J/OL]. J Immunol. 2014;192:2063–70. Article CAS Google Scholar * Kume H. RhoA/Rho-kinase as a therapeutic target in asthma[J/OL]. Curr
Med Chem. 2008;15:2876–85. Article CAS Google Scholar * Zandvakili I, Lin Y, Morris JC, Zheng Y. Rho GTPases: Anti- or pro-neoplastic targets?[J/OL]. Oncogene. 2017;36:3213–22. Article
CAS Google Scholar Download references ACKNOWLEDGEMENTS This project was supported by grants from the National Natural Science Foundation of China (#82270555, #82070538, #81870374) and
Guangdong Science and Technology Department (#2021A1515220107). AUTHOR INFORMATION Author notes * These authors contributed equally: Xiaoling Li, Mudan Zhang, Gaoshi Zhou, Shenghong Zhang.
AUTHORS AND AFFILIATIONS * Division of Gastroenterology, The First Affiliated Hospital, Sun Yat-sen University, Guangzhou, PR China Xiaoling Li, Mudan Zhang, Gaoshi Zhou, Zhuo Xie, Ying
Wang, Jing Han, Li Li, Qirui Wu & Shenghong Zhang Authors * Xiaoling Li View author publications You can also search for this author inPubMed Google Scholar * Mudan Zhang View author
publications You can also search for this author inPubMed Google Scholar * Gaoshi Zhou View author publications You can also search for this author inPubMed Google Scholar * Zhuo Xie View
author publications You can also search for this author inPubMed Google Scholar * Ying Wang View author publications You can also search for this author inPubMed Google Scholar * Jing Han
View author publications You can also search for this author inPubMed Google Scholar * Li Li View author publications You can also search for this author inPubMed Google Scholar * Qirui Wu
View author publications You can also search for this author inPubMed Google Scholar * Shenghong Zhang View author publications You can also search for this author inPubMed Google Scholar
CONTRIBUTIONS Guarantor of the article: SZ. SZ designed and oversaw the study. XL, MZ and GZ wrote and revised the manuscript. ZX, YW, JH, LL and QW revised the contents of the manuscript.
All authors approved the final manuscript and agreed to be responsible for this review. CORRESPONDING AUTHOR Correspondence to Shenghong Zhang. ETHICS DECLARATIONS COMPETING INTERESTS The
authors declare no competing interests. ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional
affiliations. RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution
and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if
changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the
material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to
obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS
ARTICLE Li, X., Zhang, M., Zhou, G. _et al._ Role of Rho GTPases in inflammatory bowel disease. _Cell Death Discov._ 9, 24 (2023). https://doi.org/10.1038/s41420-023-01329-w Download
citation * Received: 07 November 2022 * Revised: 08 January 2023 * Accepted: 11 January 2023 * Published: 23 January 2023 * DOI: https://doi.org/10.1038/s41420-023-01329-w SHARE THIS ARTICLE
Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard
Provided by the Springer Nature SharedIt content-sharing initiative
Trending News
Criminals prosecuted for drug offences after being caught with substancesThese North East criminals have been prosecuted for drug offences after being caught with illegal substances. These eigh...
Inside the newcastle corner apartment with 'stunning' cityscape viewsTake a look inside a corner apartment in Newcastle city centre, which offers 'stunning' cityscape views to pot...
How a south tyneside young women's charity is changing livesA South Tyneside based charity that works with young women across the region has helped changed the lives of over 5,000 ...
North east weather this weekend as rain is forecast after hot and dry mayThe North East weather is changing this weekend, as rain is forecast for the region after a very hot and dry May. The Me...
Tell us: Should we say goodbye to remote work?Employers certainly think so — 90% of companies want their workers back at their desks by the end of this year, accordin...
Latests News
Role of rho gtpases in inflammatory bowel diseaseABSTRACT Rat sarcoma virus homolog (Rho) guanosine triphosphatases (GTPases) function as “molecular switch” in cellular ...
Page Not Found (404) | WFAE 90.7 - Charlotte's NPR News SourceWe're sorry; the page you're looking for cannot be found. Please use the search option at the top of the page to find wh...
Sports files | phil cannon tournament director for the fedex st. Jude class | season 2013 | episode 422Sports Files Season 2013 Episode 422 | 26m 10sVideo has Closed Captions | CC Greg Gaston, TV and radio sports announcer ...
Javascript support required...
Latest relief package may not save some small businesses from bankruptcyAUDIE CORNISH, HOST: The $2 trillion spending package signed into law by President Trump at the end of December earmarke...